
Homo erectus
| Homo erectus | |
|---|---|

| |
| ’n Replika van die skedel van diePekingmensin die Paleosoölogiese Museum vanChina. | |
| Wetenskaplike klassifikasie | |
| Koninkryk: | |
| Filum: | |
| Subfilum: | |
| Klas: | |
| Orde: | |
| Suborde: | |
| Infraorde: | |
| Superfamilie: | |
| Familie: | |
| Subfamilie: | |
| Tribus: | |
| Genus: | |
| Spesie: | H. erectus
|
| Binomiale naam | |
| †Homo erectus (Dubois, 1892)
| |
| Sinonieme | |
| |
Homo erectus( "regop mens" ) is ’nuitgestorwehominienspesiewat regdeur die grootste deel van diePleistoseengeleef het. Die vroegstefossielebewyse dateer uit sowat 2 miljoen jaar gelede[1]en die mees onlangse bewyse uit sowat 108 000 jaar gelede. Die spesimens is van die eerste herkenbare lede van diegenusHomo.
Verskeie menslike spesies, soosH. heidelbergensisenH. antecessorhet blykbaar uitH. erectusontwikkel, enNeanderdallers,H. denisovaen diemoderne menshet op hulle beurt vermoedelik uitH. heidelbergensisontwikkel.[2]
H. erectuswas die eerste menslike voorouer wat deur die heleEurasiëversprei het, van dieIberiese SkiereilandtotJava.Asiatiesebevolkings vanH. erectuskon die voorouers gewees het vanH. floresiensis[3]en moontlikH. luzonensis.[4]Die laaste bekende bevolking vanH. erectusisH. e. soloensisvan Java, sowat 117 000 tot 108 000 jaar gelede.[5]
H. erectushet 'n moderner houding en liggaamsproporsies gehad en was die eerste menslike spesie met 'n plat gesig, prominente neus en moontlik 'n yl haarbedekking. Hoewel die spesie sebreinegroter as dié van sy voorouers was, het die kapasiteit grootliks gewissel na gelang van die bevolking. Dit lyk of die breine in vroeëre bevolkings vroeg in die kinderjare ophou ontwikkel het, wat daarop dui kinders was met geboorte in 'n groot mate selfonderhoudend en dat dit kognitiewe ontwikkeling deur hulle lewe beperk het.H. erectuswas 'n roofdier aan die bopunt van dievoedselketting;[6]Op terreine is die oorblyfsels ontdek van mediumgroot tot groot diere, soosbeesteofolifante.Dit lyk dus of hulle in groepe gejag het. Die spesie word verbind met die vroeë steengereedskapsbedryf en was vermoedelik die eerste menslike voorouer watvuurgebruik,[7]in groepe gejag en kos versamel het, beseerde en siek groepslede versorg het en moontlik op die see gevaar het. Hulle was dalk ook die vroegste beoefenaars vankuns,maar bewyse daarvoor is skaars.
Die mans en vroue was rofweg dieselfde grootte (dus met mingeslagsdimorfie), wat kan dui opmonogamiein ooreenstemming met algemene neigings onderprimate.Die lengte het wel van 146-185 cm gewissel en gewig van 40-68 kg. Dit is onduidelik ofH. erectuskon praat, hoewel al voorgestel is hulle het deur die een of ander prototaal gekommunikeer.
Taksonomie
[wysig|wysig bron]Ontdekking
[wysig|wysig bron]DieNederlandseanatomis Eugène Dubois en sy span het in 1891 ’n menslike kopbeen op die eiland Java in Indonesië ontdek; hy het ditPithecanthropus erectusgenoem. Die spesie is nou bekend asH. erectus.Die vonds het bekend geword as dieJavamens.Byna al die dele van die kopbeen het verlore geraak tydens dieTweede Wêreldoorlog,hoewel afgietsels nog bestaan.

Deur die20ste eeuhet antropoloë gedebatteer oor die rol vanH. erectusin die mens seevolusie.Vroeg in die eeu is vanweë nog ontdekkings op Java en by Zhoukoudian inChinageglo die moderne mens het in Asië ontwikkel. Charles Darwin en ander het egter geglo die mens se vroegste voorouers kom uit Afrika. Hy het daarop gewys datsjimpanseesengorillas,die mens se naaste lewende verwante, net in Afrika voorkom.[8]
Van die 1950's tot die 1970's het verskeie fossiele vondste in Oos-Afrika bewys gelewer dat die oudsteHomininaehier ontstaan het. Daar word nou gegloH. erectusis ’n afstammeling van vroeëre genera soosArdipithecusenAustralopithecus,of vroeëHomo-spesies soosHomo habilisofHomo ergaster.H. habilisenH. erectushet verskeie duisende jare saam bestaan en kan verskillende lyne van ’n gemeenskaplike voorouer wees.[9]
Nog fossiele is later ontdek wat eers ander name gekry het, maar later herklassifiseer is asH. erectus,soos in September 1949 inSwartkrans,Suid-Afrika,en in 1961 inTsjad,Noord-Afrika.[10]Homo erectus georgicusis die subspesienaam wat soms gebruik word vir fossiele kop- en kakebene wat in Dmanisi, Georgië, ontdek is. Hoewel dit eers as ’n aparte spesie geklassifiseer is, val dit nou binne die spesieH. erectus.
Fossiele
[wysig|wysig bron]Fossiele wat ontdek is, is onder meer:
- D2700
- KNM ER 3733
- Yuanmou-mens
- OH 9
- KNM ER 992
- KNM ER 3883
- Daka
- Sangiran 4
- Sangiran 2
- Trinil 2
- Ternifine 2-3
- Sangiran 17
- Pekingmens
- Madam Buya
- Hexian
- Ngandong 7
- Dali
- NG 6
Evolusie
[wysig|wysig bron]Daar is voorgestel datH. erectussowat 2 miljoen jaar gelede uitH. habilisontwikkel het. Dit word egter betwis, omdat hulle minstens 'n halfmiljoen jaar saam bestaan het. Alternatiewelik kon 'n groep vanH. habilisgeïsoleer gewees het en het net dié groep inH. erectusontwikkel (kladogenese).[11]
Omdat die vroegste oorblyfsels vanH. erectusin beide Afrika en Asië ontdek is (inSuid-Afrikavan sowat 2,04 miljoen jaar gelede[1][12]en inChinavan sowat 2,1 miljoen jaar gelede),[13][14][15]word gedebatteer oor waarH. erectusontwikkel het. 'n Studie van 2011 dui daarop dat ditH. habilisis wat Wes-Asië uit Afrika bereik het, dat vroeëH. erectusdaar ontwikkel het en dat vroeëH. erectustoe van Wes-Asië na Oos-Asië, Suidoos-Asië, terug na Afrika en na Europa versprei het, en eindelik in Afrika tot die moderne mens ontwikkel het.[16][17]Ander stel voorH. erectus/H. ergasterhet in Afrika ontwikkel, waar hulle eindelik in die moderne mens ontwikkel het.[18][19]
H. erectushet Sangiran, Java, teen 1,8 miljoen jaar gelede bereik,[20]en 'n tweede, aparte vlaag vanH. erectushet Zhoukoudian,China,sowat 780 000 jaar gelede beset. Vroeë tande van Sangiran is groter as en stem meer ooreen met dié vanbasaleWesterseH. erectusenH. habilisas dié wat in Zhoukoudian vanH. erectusgekry is. Latere tande van Sangiran lyk egter kleiner, wat kan dui op 'n sekondêre kolonisasievoorval in Java deur die Zhoukoudian- of 'n nabyverwante bevolking.[21]
Subspesies
[wysig|wysig bron]- Homo erectus erectus(Javamens, 1,6-0,5 miljoen jaar gelede)
- Homo erectus ergaster(1,9-1,4 miljoen jaar gelede)
- Homo erectus georgicus(Dmanisi-hominiene, 1,8-1,6 miljoen jaar gelede)
- Homo erectus lantianensis(Lantianmens, 1,6 miljoen jaar gelede)
- Homo erectus nankinensis(Nanjingmens, 0,6 miljoen jaar gelede)
- Homo erectus pekinensis(Pekingmens, 0,7 miljoen jaar gelede)
- Homo erectus soloensis(Solomens, 0,546-0,143 miljoen jaar gelede)
- Homo erectus tautavelensis(Tautavelmens, 0,45 miljoen jaar gelede)
- Homo erectus yuanmouensis(Yuanmou-mens, 1,7 miljoen jaar gelede)
Sedert die ontdekking van die Javamens in 1893 is daar 'n neiging in paleoantropologie om die getal voorgestelde spesies vanHomote verminder, en daarom sluitH. erectusalle vroeë vorme vanHomoin wat uitH. habilisontwikkel het en verskil van vroeëH. heidelbergensis(in Afrika ook bekend asH. rhodesiensis).[22]Dit word soms beskou as 'n wydverspreide, polimorfe (veelvormige) spesie.[23]
Vanweë so 'n groot variasie is al voorgestel dat die antiekeH. rudolfensisenH. habilisbeskou word as vroeë variëteite vanH. erectus.[24][25]Die primitieweH. e. georgicusvan Dmanisi, Georgië, het die kleinste breinkapasiteit van enige bekende hominien uit die Pleistoseen (sowat 600 cc), en sy insluiting in die spesie sal die omvang vanH. erectussodoende uitbrei dat dit spesies soosH. rudolfensis,H. gautengensis,H. ergasteren dalkH. habilisinsluit.[26]'n Studie uit 2015 dui egter daarop datH. georgicus'n vroeëre, primitiewer spesie vanHomoverteenwoordig.[27]Dit word soms nie asH. erectusbeskou nie.[28][29]
Daar word debatte gevoer oor of dieH. e. ergastervan Afrika 'n aparte spesie is (en datH. erectusin Asië ontwikkel en toe na Afrika migreer het),[30]en of dit die Afrikavorm (sensu lato) vanH. erectus (sensu stricto)is. As laasgenoemde reg is, is ook voorgestel datH. ergasterdie onmiddellike voorvader vanH. erectusverteenwoordig.[31]Verder is voorgestel datH. ergaster,in plaas vanH. erectusof die een of ander bastervorm tussenin, die onmiddellike voorvader van ander argaïese en moderne mense is.

Afstammelinge en sinonieme
[wysig|wysig bron]
Homo erectusis die oudste bekende spesie vanHomo.Dit het sowat 2 miljoen jaar gelede al bestaan, teenoorHomo sapiensse bykans 'n derde van 'n miljoen jaar.
Daar is geen definisie om te bepaal watter argaïese mense as 'n subspesie vanH. erectus,vanH. sapiensof as 'n aparte spesie geklassifiseer behoort te word nie.
- Kandidate virH. erectusuit Afrika:
- Homo ergaster(ofH. erectusuit Afrika ")
- Homo naledi(ofH. e. naledi)
- Kandidate virH. erectusuit Asië:
- Homo antecessor(ofH. e. antecessor)
- Homo heidelbergensis(ofH. e. heidelbergensis)
- Homo cepranensis(orH. e. cepranensis)
- Homo floresiensis[32][33]
- Homo sapiens-kandidate:
- Homo neanderthalensis(ofH. s. neanderthalensis)
- Homo denisova(ofHomosp. 'Denisova',H. sapiensssp. 'Denisova' ofH.sp. 'Altai')
- Homo rhodesiensis(ofH. s. rhodesiensis)
- Homo heidelbergensis(ofH. s. heidelbergensis)
- Homo sapiens idaltu
Anatomie
[wysig|wysig bron]Kop
[wysig|wysig bron]Homo erectushet 'n plat gesig gehad in vergelyking met vroeëre hominiene, asook 'n prominente wenkbrourif en 'n lae, plat skedel.[34][35]Die teenwoordigheid van 'n swaardlyn-, frontale en voorhoofkeel (klein kruine al langs dié beennate) het gelei tot voorstelle dat dit tekens is van 'n aansienlike verdikking van die kopbeen, veral die gewelfde skedeldak.CT-skanderingshet getoon dit is nie die geval nie. Die agterkant van die skedel is egter merkbaar dikker as by moderne mense, en is moontlik 'n basale (voorouer)-eienskap.[35][36]

Die fossielrekord toonH. erectuswas die eerste menslike spesie met 'n neus wat vooruitsteek, wat vermoedelik die gevolg is van die behoefte om klammigheid te behou wanneer droë lug ingeasem word.[37]Volgens 'n hipotese van die Amerikaanse sielkundige Lucia Jacobs was die vooruitstekende neus eerder 'n manier om die rigting van verskillende reuke vas te stel om navigasie en langafstandmigrasie te vergemaklik.[38]
Die gemiddelde breingrootte van die AsiatieseH. erectuswas sowat 1 000 cc (kubieke sentimeter). Merkbaar kleiner spesimens is ook al ontdek in Dmanisi, Georgië,Keniaen moontlikEthiopië.H. erectusse breingrootte wissel gewoonlik tussen 546 en 1 251 cc,[39]wat meer is as die wisseling by moderne mense ensjimpansees,maar minder as bygorillas.
H. erectushet die dunste tandemalje gehad van enige Plio-Pleistoseen-hominien. Emalje keer dat harde kos die tand laat breek, maar bemoeilik die skeur van taai kos. Die onderkaak byH. erectusen alle vroeëHomois dikker as dié van moderne mense en alle lewende ape. Dit beteken hulle kon baie hard byt en kou. Die praktiese implikasies daarvan is egter onduidelik. Tog was die onderkaak vanH. erectuseffens dunner as by die vroeë mens.
Die kies- en voorkiestande vanH. erectushet meer groewe as dié vanH. habilis,wat daarop duiH. erectushet brosser kos geëet (wat groewe veroorsaak). Dit alles dui daarop datH. erectusse mond minder toegerus was om harde kos te kou en beter toegerus om taai kos te skeur, maar dit het die verskeidenheid van hulle kos verklein.[40]
Liggaam
[wysig|wysig bron]
Nes die moderne mens hetH. erectusbaie in grootte gewissel, van 146 tot 185 cm in lengte en van 40 tot 68 kg in gewig. Dit was vermoedelik na aanleiding van streeksverskille in klimaat, lewensduur of voeding.[41][42]Onder primate kom dié reaksie op omgewingsfaktore net by moderne mense voor.[43][44][45]
Nes by mense, maar anders as by ander primate, was daar oënskynlik nie 'n groot grootteverskil tussen mans en vroue nie, maar daar is nie genoeg fossieldata om dit te staaf nie.[39]Breingroottes by sommige fossiele verskil wel.[39][11]

H. erectushet omtrent dieselfde lededmaatkonfigurasie en-proporsiesas moderne mense gehad. Dit dui op mensagtige beweging,[46]die eerste in dieHomo-lyn.[43]H. erectus-spore wat in Kenia ontdek is, dui ook op 'n menslike stappie.[47]'n Mensagtige skouer dui op 'n vermoë om vinnig te gooi.[48]
Dit is grootliks onduidelik wanneer menslike voorouers die meeste van hulle liggaamshare verloor het. Genetiese ontledings dui daarop dat dit min of meer ooreengestem het met toe velkleur verdonker het, sowat 1,2 miljoen jaar gelede, want die liggaam is toe blootgestel aan meer skadelikeUV-straling.[49]Dit is moontlik dat dit eers in die Pleistoseen gebeur het, omdat die groterashellingvan die aarde (wat ook dieystydperkeveroorsaak het), sou gelei het tot 'n toename in sonstraling; dit dui daarop dit het die eerste keer inAustralopithecusontstaan.[50]Australopithecushet egter blykbaar by hoër, kouer hoogtes voorgekom (gewoonlik 1 000 tot 1 600 m), waar die nagtemperatuur kan daal tot 10 of 5 °C – hulle sou dus hare nodig gehad het om warm te bly, in teenstelling met vroeëHomo,wat laer, warmer hoogtes bewoon het.[51]Bevolkings by hoërbreedtegradehet waarskynlik ligte velle ontwikkel om 'n gebrek aanvitamien Dte voorkom.[52]'nH. erectus-spesimen van 500 000 tot 300 000 jaar gelede vanTurkyeis gediagnoseer met die eerste voorval van 'n siekte weens 'n vitamien D-gebrek wat gewoonlik onder donkervellige mense by hoë breedtegrade voorkom.[53]
Daar word geglo haarloosheid het sweet vergemaklik,[54]maar 'n afname inparasieteenseksuele seleksieis ook al voorgestel.[55][56]

Metabolisme
[wysig|wysig bron]
Die spesimen van die 1,8 miljoen jaar oueMojokerto-kind,wat op sowat een jaar oud dood is, het sowat 72 tot 84% van die gemiddelde volwasse breingrootte gehad, wat eerder ooreenstem met die vinnige groei van die breine van die groot ape as van mense. Dit dui daarop datH. erectuswaarskynlik nie kognitief was in vergelyking met moderne mense nie en dat die lang kinderlewe van mense omdat die brein lank vat om volgroei te raak, eers later ontwikkel het – dalk in die mens/Neanderdaller semees onlangse gemeenskaplike voorsaat.[57]
Gebaseer op 'n gemiddelde massa van 63 kg vir mans en 52,3 kg vir vroue, sou die daaglikse energiegebruik (die getal kalorieë per dag gebruik) onderskeidelik na raming sowat 2 271,8 en 1 909,5 gewees het. Dit is soortgelyk as by die vroeëHomo,ondanks 'n merkbare toename in aktiwiteits- en migrasiekapasiteit.
Dit is moontlik omdat die langer bene vanH. erectusmeer energiedoeltreffend oor lang afstande was. Nogtans is die raming virH. erectus-vroue 84% hoër as byAustralopithecus-vroue, waarskynlik vanweë 'n groter liggaamsgrootte en laer groeitempo.[58]
Beendikte
[wysig|wysig bron]
Die buitenste laag van diebeenis buitengewoon verdik, veral in Oos-Asiatiese bevolkings. Die murgkanaal in die lang bene (waar diebeenmurggeberg word) is uiters vernou. Dié graad van verdikking word gewoonlik aangetref by halfseediere wat hulle swaar bene gebruik om hulle te help sink. Manlike spesimens het dikker buitelae as vroue.[59]
Voordat vollediger skelette vanH. erectusontdek is, is voorgestel lede van die spesie was reusagtig en dat die verdikte bene nodig was om hulle groot massa te steun. Daar was 'n hipotese dat die intense fisieke aktiwiteit tot die verdikking van die bene kon gelei het, maar in 1970 het die bioloog Stanley Marion Garn gedemonstreer daar is min verband tussen die twee, of minstens in moderne mense. Garn het wel opgelet daar is 'n groter verskil in die dikte van die buitelaag tussen verskillende bevolkingsgroepe, en het afgelei dit is vanweë genetiese eerder as omgewingsfaktore.
In 2004 het Noel Boaz en Russel Ciochon voorgestel dit was die gevolg van 'n kultuurpraktyk, waarinH. erectusmet vuiste, stene en knuppels teen mekaar sou veg om dispute op te los of vir maats te baklei, want die kopbeen is op sleutelplekke verdik. Die kakebeen is taamlik robuus en kan harde houe weerstaan; die swaar wenkbrourif beskerm die oë en harde been bedek die ore en loop tot agter in die kop, wat beteken houe teen enige van dié dele kan doeltreffend oor die hele kopbeen versprei word; ook die bokant van die skedel, waar die brein is, word deur harde been beskerm. Baie fossiele skedels toon breuke aan die bokant, maar hulle kan ook tekens van herstel en oorlewing toon. Die antropoloog Peter Brown stel voor daar is 'n soortgelyke rede dat die moderne Australieseaboriginesse kopbeen ongewoon dik is: Dit is vanweë 'n ritueel wat in Sentraal- en Suidoos-Australiese stamme gewild is waar teenstanders mekaar met stokke sal slaan tot een bewusteloos is.[60]
Kultuur
[wysig|wysig bron]Sosiale struktuur
[wysig|wysig bron]
Die enigste fossiele getuienis vir die samestelling vanH. erectus-groepe kom van vier terreine buite Ileret, Kenia, waar 97 voetspore sowat 1,5 miljoen jaar gelede getrap is deur 'n groep van minstens 20 individue. Gebaseer op die grootte van die versteende voetspore, kon een van die paadjies getrap gewees het deur 'n groep mans, wat daarop dui dat hulle 'n gespesialiseerde taak onderneem het, soos om te jag, kos te versamel of op 'n grenspatrollie te gaan. As dit korrek is, dui dit op 'n verdeling van arbeid volgens geslagte, wat menslike gemeenskappe onderskei van dié van ander groot ape en sosiale, vleisvretende soogdiere. In moderne gemeenskappe vanjagter-versamelaarswat groot prooi jag, gaan mans gewoonlik op veldtogte om groot diere te jag, terwyl vroue op voorspelbaarder kosse fokus.[47]
Gebaseer op die gedrag van moderne savannesjimpansees en-bobbejane,hetH. erectus ergasterdalk in groot groepe met baie mans gewoon om die gemeenskap teen groot roofdiere in die oop en blootgesteldesavanneste verdedig.[61]Verspreidingspatrone dui egter daarop datH. erectusgewoonlik gebiede met baie vleisvreters vermy het.[62]Dit is moontlik dat die vriendskappe tussen mans en tussen vroue 'n belangrikse sosiale aspek was.[61]
OmdatH. erectus-kinders vinniger breingroeitempo's gehad het, is waarskynlik nie so baie tyd aan kinders grootmaak bestee soos by moderne mense nie.[63]
Omdat diegeslagsdimorfietussen mans en vroue klein was, word algemeen aanvaar hulle was monogaam, want kleiner geslagsdimorfie in primate is gewoonlik 'n aanduiding van dié paringstelsel.[64]Dit is egter onduidelik ofH. erectuswel dieselfde lae voorkoms van geslagsdimorfie gehad het.[11]As dit die geval is, beteken dit net vroue se lengte het toegeneem sedert die voorouerspesie, en dit kon veroorsaak gewees het deur 'n verandering in vroulike vrugbaarheid of dieet en/of 'n afname in druk op mans om groter te wees. Dit kan weer dui op 'n verandering in vroue se gedrag wat dit moeiliker vir mans sou gemaak het om 'n harem te onderhou.[65]
Voedsel
[wysig|wysig bron]'n Toename in breingrootte word dikwels verbind met meer vleis in die dieet en dus 'n groter kalorie-inname. 'n Toename in proteïeninname deur insekte is ook al voorgestel as 'n oorsaak. Dit is egter ook moontlik dat die energiekosbare binnegoed inH. erectuskleiner geword het omdat die binnegoed van die groter ape gebruik word om vet te sintetiseer deur plantmateriaal te laat gis, en dit is byH. erectusvervang deur dieetkundige diervet, wat toegelaat het dat meer energie vir breingroei gebruik word. Dit sou breingrootte indirek vergroot het terwyl dieselfde kalorievereistes van voorouerspesies gehandhaaf is.H. erectuskon ook die eerste spesie gewees het wat 'njag-en-versamel-tegniek ontwikkel het in reaksie op 'n groter afhanklikheid van vleis. Met 'n klem op spanwerk, verdeling van arbeid en die deel van voedsel was jag-en-versamel 'n drasties verskillende voedselstrategie as vroeër.[40][66]

Terreine vanH. erectusword dikwels verbind met oorblyfsels van mediumgroot tot groot wilde diere, soosolifante,renosters,seekoeie,beesteenbere.Hulle sou baie oorskietkos gehad het, en dit dui op die moontlike deel van kos of langtermynpreservering (soos die uitdroging van kos) as hulle die hele prooi wou gebruik. Dit is moontlik datH. erectusafhanklik van die vleis van groot diere geraak het, en die verdwyning vanH. erectusuit dieLevantword verbind met die plaaslike uitsterwing van die Europese wilde-olifant (met sy reguit slagtande).[66]
H. erectusse dieet het egter grootliks verskil na gelang van gebied. In Israel is byvoorbeeld 'n terrein ontdek waar die inwoners 55 verskillende soortevrugte,groente,sade,neuteenknolleversamel en geëet het, en dit lyk of hulle sekere plantemateriale eers op 'n vuur gebraai het; hulle het ookamfibieë,reptiele,voëlsen andergewerweldesgeëet saam met die gewone diere soos olifante endamherte.[67]In Kenia is 'n terrein ontdek waar inwoners (saam met die gewone beeste, renosters en seekoeie) waterdiere soosseeskilpaaie,krokodilleenkatvissegeëet het. Die aas van groot diere is moontlik op die terrein geëet, maar die skilpaaie en katvisse is waarskynlik lewend versamel.[68]
H. erectusse monde en tande was nie so veelsydig as dié van voorouerspesies nie en hulle kon dus 'n kleiner verskeidenheid kos eet. Gereedskap is egter waarskynlik gebruik om harde kos te verwerk en dit het die gebit beïnvloed. Dié kombinasie het hulle dalk 'n dieetkundige veelsydigheid gegee. Dit hetH. erectusmoontlik toegelaat om 'n verskeidenheid omgewings te bewoon en na buite Afrika te migreer.[40]
Tegnologie
[wysig|wysig bron]Gereedskapvervaardiging
[wysig|wysig bron]

H. erectuskry die krediet daarvoor dat hy dieAcheuleen-steengereedskapbedryf ontdek het, wat die Oldowanbedryf voorafgegaan het,[69][70]en was die eerste beskawing wat skerp, tweesydige handbyle gemaak het.[71][72]Hulle was waarskynlik veeldoelige instrumente wat vir verskeie soorte aktiwiteit gebruik is, soos om vleis, hout of eetbare plante te sny.[73]
Die vroegste rekord van Acheuleentegnologie kom van Wes-Turkana, Kenia, van 1,76 miljoen jaar gelede. Die Acheuleen was die tweede vroegste tydvak van diePaleolitikum.[74]Oldowangereedskap is ook op die terrein ontdek, en die twee tegnologieë het blykbaar vir 'n ruk saam voorgekom. Die vroegste rekords van die Acheuleentegnologie buite Afrika dateer van nie vroeër as 'n miljoen jaar gelede nie, en dit dui daarop dat dit eers wydverspreid geword het ná 'n sekondêre verspreiding vanH. erectusuit Afrika.[72]
Op Java hetH. erectusgereedskap van skulpe gemaak by Sangiran[75]en Trinil.[76]Ronde klippe met 'n deursnee van 6 tot 12 cm word dikwels aangetref op terreine in Afrika en China uit die vroeë Paleolitikum.[77]
Op terreine in Oos-Afrika, soos in Kenia, is moontlike bewyse gevind dat vroeë mense vuur gebruik het. Die herverhitting van sekere stukke klei van 1,42 miljoen jaar oud[78]het gewys die klei moes tot 400 ºC verhit gewees het om te verhard. Geen bewyse is egter nog gevind dat hy sy kos gaargemaak het nie.
Konstruksie
[wysig|wysig bron]
In 1962 is 'n sirkel van 366 × 427 × 30 cm in Olduvai Gorge ontdek wat van vulkaanrots gemaak is. Elke 61 tot 76 cm is rotse 15 tot 23 cm hoog opmekaargestapel. Die Britse paleoantropoloog Mary Leakey het voorgestel die rotshope is gebruik om pale te ondersteun wat in die grond gedruk is, moontlik om 'n windskuiling of rowwe hut te bou. Sommige modernenomadiese stammebou soortgelyke lae rotsmure as tydelike beskuttings; hulle buig reguit takke as pale en gebruik gras of diervel as bedekking.[80]Dit dateer van 1,75 miljoen jaar gelede en is die oudste beweerde voorbeeld vanargitektuur.[81]
In Europa is glo bewyse van geboude skuilings wat tot 424 000 jaar gelede terugstrek, ontdek in Bilzingsleben, Duitsland; Terra Amata, Frankryk; en Fermanville en Saint-Germain-des-Vaux inNormandië.Die oudste bewys van 'n skuiling (en kampvuur) in Europa kom uit Přezletice,Tsjeggië;dit dateer uit 700 000 jaar gelede. Dié skuiling se basis is sowat 3 x 4 m aan die buitekant en 3 x 2 m aan die binnekant, en was moontlik 'n ferm hut wat as winterbasis gebruik is.[82]
Die vroegste bewyse van grotbewoning is ontdek in dieWonderwerkgrot,Suid-Afrika.Dit dateer van sowat 1,6 miljoen jaar gelede. Die gebruik van grotte wêreldwyd is egter sporadies tot omtrent 600 000 jaar gelede.[83]
Klere
[wysig|wysig bron]
Dit is onduidelik of klere al ontwerp was. Die vroegste raming daarvoor is sowat 3 miljoen jaar gelede as gevolg van 'n gebrek aan isolerende liggaamshare.[50]Dit is bekend dathaar- en liggaamsluiseby moderne mense sowat 170 000 jaar gelede van mekaar begin verskil het (liggaamsluise kom net voor op mense wat klere dra). Dit was lank voordat die moderne mens uit Afrika getrek het en dit beteken klere is gedra lank voordat hulle in kouer klimaatstreke gaan woon het.
Een van die eerste gebruike vir diervelle is vermoedelik as klere, en die eerste velverwerking het sowat 780 000 jaar gelede voorgekom, hoewel dit nie noodwendig op klere dui nie.[84]
Seevaart
[wysig|wysig bron]Acheuleen-artifakte van die Pleistoseen wat op afgeleë eilande ontdek is en wat nooit op land aangetref is nie, wys dalk datH. erectusdie see so vroeg as 'n miljoen jaar gelede inIndonesiëbevaar het. Hulle het op die eilandeFlores,TimorenRotiaangekom, wat beteken hulle moes dieLombokstraatoorgesteek het, minstens voor 800 000 jaar gelede. Dit is moontlik dat hulle ook die eerste Europese matrose was en dat hulle dieStraat van GibraltartussenNoord-AfrikaenSpanjeoorgesteek het. 'n Genetiese ontleding in 2021 van dié eilande se bevolkings vanH. erectuskon nie tekens vind van verbastering met moderne mense nie.[85]Seevaartvermoëns sal bewysH. erectuskon vooruit beplan, dalk soms maande voor 'n seereis.[86][87]
Net so dateerHomo luzonensisvan tussen 771 000 en 631 000 jaar gelede. OmdatLuzonin dieKwartêraltyd 'n eiland was, souH. luzonensisse voorouers 'n groot seekruising moes onderneem het om daar te kom.[88]
Gesondheidsorg
[wysig|wysig bron]
Moontlik die eerste tekens van 'n siek groepslid is 'n 1,77 miljoen jaar oue spesimen vanH. e. georgicuswat al sy tande behalwe een verloor het weens ouderdom of 'n tandvleissiekte. Dit is die eerste voorbeeld van iemand wie se kouvermoë kwaai aangetas is, maar hy het jare daarna nog oorleef. Dit is egter moontlik dat Australopithecus siek groepslede kon versorg het.[89]
Omdat diéH. e. georgicusnie kon kou nie, het hy waarskynlik sagte plant- of dierkos geëet, dalk met die hulp van ander groepslede. Groepe wat by hoë breedtegrade gewoon het, was vermoedelik hoofsaaklik vleisetend. Hulle sou dus onder meer sagte weefsel soos beenmurg of breine geëet het, wat individue sonder tande sou help oorleef het.[90]
Die 1,5 miljoen jaar oueTurkana-seunis gediagnoseer met 'n rugwerwelbesering en omdat hy steeds gegroei het, het dit 'n abnormale kurwe in die rug veroorsaak. Dit sou laerugpyn veroorsaak het en loop, buig en ander daaglikse bedrywighede belemmer het. Dit lyk of hy tot in sy laatadolessensieoorleef het, en dit dui op gevorderde groepsorg.[91]
Kuns en rituele
[wysig|wysig bron]

'n Gegraveerde skulp met geometriese merke kan dalk 'n bewys wees van die vroegte kuns. Dit dateer van 546 000 tot 436 000 jaar gelede. Die vermoë om kuns te vervaardig kan 'n aanduiding wees van 'n simboliese denkvermoë, wat verbind word met modernekognisieen gedrag.[76][92]
In 1976 het die Amerikaanse argeoloog Alexander Marshack gesê gegraveerde lyne op 'n osribbebeen van Pech de l'Azé, Frankryk, wat met die Acheuleen verbind word, is soortgelyk aan eenvoudige ontwerpe wat in Paleolitiese grotontwerpe van moderne mense voorkom.[93]Drie krale van die eierdoppe vanvolstruisewat met die Achueleen verbind word, is ontdek in Noordwes-Afrika. Dit was die oudste krale wat nog ontdek is. Krale uit die Acheuleen is ook in Frankryk en Israel ontdek.[86]
Die "Venus van Tan-Tan" vanMarokkoen "Venus van Berekhat Ram" van dieGolanhooglanduit die Middel-Pleistoseen is vermoedelik deurH. erectusgeskep om 'n menslike vorm te verteenwoordig. Hulle is hoofsaaklik deur natuurlike verwering gevorm, maar effens verander om klem op sekere groewe te lê om soos haarlyne, ledemate en oë te lyk.[94][95]Eersgenoemde "Venus" het tekens van pigment aan die voorkant, wat moontlik 'n aanduiding van kleuring is.[94]
H. erectuswas ook die eerste mens wat rooi pigment,oker,doelbewus versamel het, van so vroeg as die Middel-Pleistoseen. Okerklonte by Olduvaikloof inTanzanië,wat met die 1,4 miljoen jaar oue Olduvai-hominied 9 verbind word, en Ambrona, Spanje – onderskeidelik van 424 000 tot 374 000 jaar gelede – is vermoedelik met 'n hamersteen geslaan en doelbewus gevorm.[96][93]By Terra Amata, Frankryk, wat van sowat 400 000 jaar gelede dateer, is geel en bruin oker ontdek en dit word verbind met paalstrukture. Oker is waarskynlik verhit om dié kleure te verkry.[96][97]Omdat dit onduidelik is ofH. erectusoker kon gebruik het vir enige praktiese aanwending, kan die versameling van die oker daarop dui datH. erectusdie vroegste mense was wat 'n sin virestetikagehad het en dat hulle verder as net aan oorlewing gedink het. Dit is egter onduidelik of dié paar voorbeelde nie net afgesonderde gebruike van oker was nie, want oker kom meer dikwels voor op terreine van die Middel- en Laat Pleistoseen wat verbind word met die Neanderdallers enH. sapiens.[98][93]
Taal
[wysig|wysig bron]In 1984 het die werwelkolom van die 1,6 miljoen jaar oue adolessent die Turkana-seun aangedui hy het nie die nodige asemhalingsorgane gehad om te kon praat nie. In 2001 het die Amerikaanse antropoloë Bruce Latimer en James Ohman tot die gevolgtrekking gekom die seun het aan werwelkolomafwykings of-beseringsgely.[99]In 2006 het die Amerikaanse antropoloog Marc Meyer en sy kollegas 'nH. e. georgicus-spesimen van 1,8 miljoen jaar gelede beskryf wat 'nruggraatgehad het wat binne die omvang van variëteit van menslike ruggrate geval het. Hulle het tot die gevolgtrekking gekom dat die Turkana-seun se ruggraat 'n afwyking is en nie die spesie verteenwoordig nie. Hulle het verder gegloH. e. georgicuswas die voorouer van alleH. erectuswat nie uit Afrika was nie, en dat die asemhalingspiere van alleH. erectus(of minstens dié wat nieH. ergasterwas nie) nie die spraakvermoë sou aangetas het nie.[100]
Neurologies het alleHomo-spesies breine wat eenders gevorm is, en daarom was die streke van die brein wat sinformulering en spraak inHomo erectusbeheer het, vergelykbaar met dié van moderne mense. Dit is egter nie noodwendig 'n aanduiding van spraak nie, want dieselfde streke in 'nsjimpanseese brein gee hom nie die vermoë om te kan praat nie.[101]
'n Been wat die tong beheer uit 'nH. erectus-fossiel van 400 000 jaar gelede van Castel di Guido,Italië,dui daarop datH. erectusnie 'n menslike stemorgaan en dus nie die voorvereistes vir menslike spraak gehad het nie.[102]
Uitsterwing
[wysig|wysig bron]Die laaste bekende voorkoms vanHomo erectusis volgens 'n studie wat in 2019 gepubliseer is, sowat 117 000 tot 108 000 jaar gelede in Ngandong, Java.[5]
In 2020 het navorsers berigHomo erectusenHomo heidelbergensishet net voor hulle uitsterwing meer as die helfte van hulle klimaatsnis (klimaat waaraan hulle gewoond was) verloor sonder enige fisieke aanpassing, en dat klimaatsverandering 'n aansienlike rol gespeel het in die uitsterwing van vorigeHomo-spesies.[103][104][105]
Filogenie
[wysig|wysig bron]'n Kladogram vanHomo erectuslyk soos volg[106](met 'n aanduiding van hoeveel miljoen jaar gelede die klades gesplits het):
| Homo(2,85) |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Homo erectuswas oorspronklik van Afrika. Die oorgebleweHomo heidelbergensis(wat kladistiesHomo sapiensen oorspronklik van Afrika was), het binne die AsiatieseHomo erectusverskyn. Dit lyk of kontemporêre groepe verbaster het, en enige filogenie soos dié gee net 'n rowwe indruk van die evolusie vanHomo:Uitgestorwe lyne kon gedeeltelik in ander groepe voortbestaan het. Nie ingesluit nie, is ander kontemporêre groepe soosHomo floresiensis,Homo naledi,Homo luzonensis,Homo rudolfensis,Australopithecus sediba,Australopithecus africanusenParanthropus.
Galery
[wysig|wysig bron]-
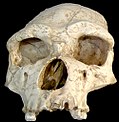 ’n Kopbeen vanHomo erectus tautavelensis.
’n Kopbeen vanHomo erectus tautavelensis. -
 Replika van die onderste kakebeen vanHomo erectus tautavelensis.
Replika van die onderste kakebeen vanHomo erectus tautavelensis. -
 Die Calvaria-kopbeen.
Die Calvaria-kopbeen. -
 ’n Rekonstruksie gebaseer op die Daka-kopbeen, Ethiopië.
’n Rekonstruksie gebaseer op die Daka-kopbeen, Ethiopië. -
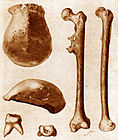 Pithecanthropus erectus-bene (nouH. erectus), in 1891 in Java ontdek.
Pithecanthropus erectus-bene (nouH. erectus), in 1891 in Java ontdek.





Verwysings
[wysig|wysig bron]- ↑1,01,1Herries AI, Martin JM, Leece AB, Adams JW, Boschian G, Joannes-Boyau R, et al. (April 2020)."Contemporaneity ofAustralopithecus,Paranthropus,and earlyHomo erectusin South Africa ".Science.368(6486): eaaw7293.doi:10.1126/science.aaw7293.PMID32241925.
- ↑Dembo M, Radovčić D, Garvin HM, Laird MF, Schroeder L, Scott JE, et al. (Augustus 2016). "The evolutionary relationships and age of Homo naledi: An assessment using dated Bayesian phylogenetic methods".Journal of Human Evolution.97:17–26.doi:10.1016/j.jhevol.2016.04.008.hdl:2164/8796.PMID27457542.
- ↑van den Bergh GD, Kaifu Y, Kurniawan I, Kono RT, Brumm A, Setiyabudi E, et al. (Junie 2016). "Homo floresiensis-like fossils from the early Middle Pleistocene of Flores".Nature.534(7606): 245–248.Bibcode:2016Natur.534..245V.doi:10.1038/nature17999.PMID27279221.S2CID205249218.
- ↑Détroit F, Mijares AS, Corny J, Daver G, Zanolli C, Dizon E, et al. (April 2019)."A new species of Homo from the Late Pleistocene of the Philippines"(PDF).Nature.568(7751): 181–186.Bibcode:2019Natur.568..181D.doi:10.1038/s41586-019-1067-9.PMID30971845.S2CID106411053.
- ↑5,05,1Rizal Y, Westaway KE, Zaim Y, van den Bergh GD, Bettis EA, Morwood MJ, et al. (Januarie 2020). "Last appearance of Homo erectus at Ngandong, Java, 117,000-108,000 years ago".Nature.577(7790): 381–385.doi:10.1038/s41586-019-1863-2.PMID31853068.S2CID209410644.
- ↑Ben-Dor M, Sirtoli R, Barkai R (Augustus 2021)."The evolution of the human trophic level during the Pleistocene".American Journal of Physical Anthropology.175(Suppl 72): 27–56.doi:10.1002/ajpa.24247.PMID33675083.
- ↑Zohar, Irit; Alperson-Afil, Nira; Goren-Inbar, Naama; Prévost, Marion; Tütken, Thomas; Sisma-Ventura, Guy; Hershkovitz, Israel; Najorka, Jens (14 November 2022)."Evidence for the cooking of fish 780,000 years ago at Gesher Benot Ya'aqov, Israel".Nature Ecology & Evolution(in Engels).6(12): 2016–2028.doi:10.1038/s41559-022-01910-z.ISSN2397-334X.
- ↑Darwin, Charles R. (1871).The Descent of Man and Selection in Relation to Sex.John Murray.ISBN0-8014-2085-7.
- ↑F. Spoor, M. G. Leakey, P. N. Gathogo, F. H. Brown, S. C. Antón, I. McDougall, C. Kiarie, F. K. Manthi & L. N. Leakey (9 Augustus 2007). "Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya".Nature.448(7154): 688–691.doi:10.1038/nature05986.
{{cite journal}}:AS1-onderhoud: meer as een naam (link) - ↑Kalb, Jon E (2001).Adventures in the Bone Trade: The Race to Discover Human Ancestors in Ethiopia's Afar Depression.Springer. p. 76.ISBN0-387-98742-8.Besoek op2 Desember2010.
- ↑11,011,111,2Spoor F, Leakey MG, Gathogo PN, Brown FH, Antón SC, McDougall I, et al. (Augustus 2007). "Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya".Nature.448(7154): 688–691.Bibcode:2007Natur.448..688S.doi:10.1038/nature05986.PMID17687323.S2CID35845.
- ↑"Our direct human ancestor Homo erectus is older than we thought".EurekAlert.AAAS.
- ↑Zhu Z, Dennell R, Huang W, Wu Y, Qiu S, Yang S, et al. (Julie 2018). "Hominin occupation of the Chinese Loess Plateau since about 2.1 million years ago".Nature.559(7715): 608–612.Bibcode:2018Natur.559..608Z.doi:10.1038/s41586-018-0299-4.PMID29995848.S2CID49670311.
- ↑Barras C (2018)."Tools from China are oldest hint of human lineage outside Africa".Nature.doi:10.1038/d41586-018-05696-8.ISSN0028-0836.S2CID188286436.
- ↑Hao L, Chao Rong L, Kuman K (2017). "Longgudong, an Early Pleistocene site in Jianshi, South China, with stratigraphic association of human teeth and lithics".Science China Earth.60(3): 452–462.Bibcode:2017ScChD..60..452L.doi:10.1007/s11430-016-0181-1.S2CID132479732.
- ↑Ferring R, Oms O, Agustí J, Berna F, Nioradze M, Shelia T, et al. (Junie 2011)."Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85-1.78 Ma".Proceedings of the National Academy of Sciences of the United States of America.108(26): 10432–10436.Bibcode:2011PNAS..10810432F.doi:10.1073/pnas.1106638108.PMC3127884.PMID21646521.
- ↑Augusti J, Lordkipanidze D (Junie 2011). "How" African "was the early human dispersal out of Africa?".Quaternary Science Reviews.30(11–12): 1338–1342.Bibcode:2011QSRv...30.1338A.doi:10.1016/j.quascirev.2010.04.012.
- ↑Rightmire GP (1998). "Human Evolution in the Middle Pleistocene: The Role ofHomo heidelbergensis".Evolutionary Anthropology.6(6): 218–227.doi:10.1002/(sici)1520-6505(1998)6:6<218::aid-evan4>3.0.co;2-6.S2CID26701026.
- ↑Asfaw B, Gilbert WH, Beyene Y, Hart WK, Renne PR, WoldeGabriel G, et al. (Maart 2002). "Remains of Homo erectus from Bouri, Middle Awash, Ethiopia".Nature.416(6878): 317–320.Bibcode:2002Natur.416..317A.doi:10.1038/416317a.PMID11907576.S2CID4432263.
- ↑Husson, Laurent; Salles, Tristan; Lebatard, Anne-Elisabeth; Zerathe, Swann; Braucher, Régis; Noerwidi, Sofwan; Aribowo, Sonny; Mallard, Claire; Carcaillet, Julien; Natawidjaja, Danny H.; Bourlès, Didier; ASTER team; Aumaitre, Georges; Bourlès, Didier; Keddadouche, Karim (8 November 2022). "Javanese Homo erectus on the move in SE Asia circa 1.8 Ma".Scientific Reports(in Engels).12(1): 19012.Bibcode:2022NatSR..1219012H.doi:10.1038/s41598-022-23206-9.ISSN2045-2322.
- ↑Zaim Y, Ciochon RL, Polanski JM, Grine FE, Bettis EA, Rizal Y, et al. (Oktober 2011). "New 1.5 million-year-old Homo erectus maxilla from Sangiran (Central Java, Indonesia)".Journal of Human Evolution.61(4): 363–376.doi:10.1016/j.jhevol.2011.04.009.PMID21783226.
- ↑Kaifu Y, Baba H, Aziz F, Indriati E, Schrenk F, Jacob T (Desember 2005). "Taxonomic affinities and evolutionary history of the Early Pleistocene hominids of Java: dentognathic evidence".American Journal of Physical Anthropology.128(4): 709–726.doi:10.1002/ajpa.10425.PMID15761880.
- ↑Perkins S (2013). "Skull suggests three early human species were one".Nature.doi:10.1038/nature.2013.13972.S2CID88314849.
- ↑Lordkipanidze D, Ponce de León MS, Margvelashvili A, Rak Y, Rightmire GP, Vekua A, Zollikofer CP (Oktober 2013). "A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo".Science.342(6156): 326–331.Bibcode:2013Sci...342..326L.doi:10.1126/science.1238484.PMID24136960.S2CID20435482.
- ↑Black R (17 Oktober 2013)."Beautiful Skull Spurs Debate on Human History".National Geographic.Besoek op6 Junie2021.
- ↑Sample I (17 Oktober 2013)."Skull of Homo erectus throws story of human evolution into disarray".The Guardian.
- ↑Giumares SW, Merino CL (September 2015)."Dmanisi hominin fossils and the problem of multiple species in the early Homo genus"(PDF).Nexus: The Canadian Student Journal of Anthropology.23.S2CID73528018.Geargiveer vanafdie oorspronklike(PDF)op 14 Januarie 2020.
- ↑Argue D, Groves CP, Lee MS, Jungers WL (Junie 2017). "The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters".Journal of Human Evolution.107:107–133.doi:10.1016/j.jhevol.2017.02.006.PMID28438318.
- ↑The International Encyclopedia of Biological Anthropology.(2018-10-04). John Wiley & Sons, Inc.. 1–4.DOI:10.1002/9781118584538.ieba0139.ISBN 9781118584422.
- ↑Baab K (Desember 2015)."Defining Homo erectus".Handbook of Paleoanthropology(2 uitg.): 2189–2219.doi:10.1007/978-3-642-39979-4_73.ISBN978-3-642-39978-7.
- ↑Tattersall I, Schwartz J (2001).Extinct Humans.Boulder, Colorado: Westview/Perseus.ISBN978-0-8133-3482-0.
- ↑In 2017 is voorgestelH. floresiensisis vanweëmorfologieseoorwegings 'n susterspesie van ófH. habilisóf 'nhabilis-erectus-ergaster-sapiens-klade,en is 'n veel antieker lyn asHomo erectusself.
- ↑Argue D, Groves CP, Lee MS, Jungers WL (Junie 2017). "The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters".Journal of Human Evolution.107:107–133.doi:10.1016/j.jhevol.2017.02.006.PMID28438318.
- ↑Baba H, Aziz F, Kaifu Y, Suwa G, Kono RT, Jacob T (Februarie 2003). "Homo erectus calvarium from the Pleistocene of Java".Science.299(5611): 1384–1388.doi:10.1126/science.1081676.PMID12610302.S2CID20437090.
- ↑35,035,1Balzeau A (2006)."Are thickened cranial bones and equal participation of the three structural bone layers autapomorphic traits ofHomo erectus?".Bulletins et mémoires de la Société d'Anthropologie de Paris.18(3–4): 145–163.doi:10.4000/bmsap.1528.
- ↑Copes LE, Kimbel WH (Januarie 2016)."Cranial vault thickness in primates: Homo erectus does not have uniquely thick vault bones".Journal of Human Evolution.90:120–134.doi:10.1016/j.jhevol.2015.08.008.PMID26767964.
- ↑Franciscus RG, Trinkaus E (April 1988). "Nasal morphology and the emergence of Homo erectus".American Journal of Physical Anthropology.75(4): 517–527.doi:10.1002/ajpa.1330750409.PMID3133950.
- ↑Jacobs LF (Februarie 2019)."The navigational nose: a new hypothesis for the function of the human external pyramid".The Journal of Experimental Biology.222(Pt Suppl 1): jeb186924.doi:10.1242/jeb.186924.PMID30728230.
- ↑39,039,139,2Antón SC, Taboada HG, Middleton ER, Rainwater CW, Taylor AB, Turner TR, et al. (Julie 2016)."Morphological variation in Homo erectus and the origins of developmental plasticity".Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences.371(1698): 20150236.doi:10.1098/rstb.2015.0236.PMC4920293.PMID27298467.
- ↑40,040,140,2Ungar PS, Grine FE (2006). "Diet in EarlyHomo:A Review of the Evidence and a New Model of Adaptive Versatility ".Annual Review of Anthropology.35:208–228.doi:10.1146/annurev.anthro.35.081705.123153.
- ↑Alan Walker, Richard Leakey (1993).The Nariokotome Homo erectus skeleton.Harvard University Press. p. 412.ISBN9780674600751.Besoek op2 Oktober2022.
- ↑Migliano AB, Guillon M (2012)."The Effects of Mortality, Subsistence, and Ecology on Human Adult Height and Implications forHomoEvolution ".Current Anthropology.53(S6): 359–368.doi:10.1086/667694.S2CID84442763.
- ↑43,043,1Antón, Susan C.; Taboada, Hannah G.; Middleton, Emily R.; Rainwater, Christopher W.; Taylor, Andrea B.; Turner, Trudy R.; Turnquist, Jean E.; Weinstein, Karen J.; Williams, Scott A. (5 Julie 2016)."Morphological variation in Homo erectus and the origins of developmental plasticity".Philosophical Transactions of the Royal Society B: Biological Sciences.371(1698): 20150236.doi:10.1098/rstb.2015.0236.ISSN0962-8436.PMC4920293.PMID27298467.
- ↑Antón, Susan C.; Potts, Richard; Aiello, Leslie C. (4 Julie 2014)."Evolution of earlyHomo:An integrated biological perspective ".Science(in Engels).345(6192): 1236828.doi:10.1126/science.1236828.ISSN0036-8075.PMID24994657.S2CID30188239.
- ↑Antón, Susan C.; Taboada, Hannah G.; Middleton, Emily R.; Rainwater, Christopher W.; Taylor, Andrea B.; Turner, Trudy R.; Turnquist, Jean E.; Weinstein, Karen J.; Williams, Scott A. (5 Julie 2016)."Morphological variation inHomo erectusand the origins of developmental plasticity ".Philosophical Transactions of the Royal Society B: Biological Sciences.371(1698): 20150236.doi:10.1098/rstb.2015.0236.ISSN0962-8436.PMC4920293.PMID27298467.
- ↑Ruff C (Maart 2008). "Femoral/humeral strength in early African Homo erectus".Journal of Human Evolution.54(3): 383–390.doi:10.1016/j.jhevol.2007.09.001.PMID17977577.
- ↑47,047,1Hatala KG, Roach NT, Ostrofsky KR, Wunderlich RE, Dingwall HL, Villmoare BA, et al. (Julie 2016)."Footprints reveal direct evidence of group behavior and locomotion in Homo erectus".Scientific Reports.6(28766): 28766.Bibcode:2016NatSR...628766H.doi:10.1038/srep28766.PMC4941528.PMID27403790.
- ↑Roach, & Richmond. (2015). "Clavicle length, throwing performance and the reconstruction of the Homo erectus shoulder".Journal of Human Evolution,80(C), 107–113.
- ↑Rogers AR, Iltis D, Wooding S (2004). "Genetic Variation at the MC1R Locus and the Time since Loss of Human Body Hair".Current Anthropology.45(1): 105–108.doi:10.1086/381006.S2CID224795768.
- ↑50,050,1Gilligan I (2010). "The Prehistoric Development of Clothing: Archaeological Implications of a Thermal Model".Journal of Archaeological Method and Theory.15:15–80.doi:10.1007/s10816-009-9076-x.S2CID143004288.
- ↑Dávid-Barrett T, Dunbar RI (Mei 2016)."Bipedality and hair loss in human evolution revisited: The impact of altitude and activity scheduling".Journal of Human Evolution.94:72–82.doi:10.1016/j.jhevol.2016.02.006.PMC4874949.PMID27178459.
- ↑Jablonski NG (Maart 2012). "Human skin pigmentation as an example of adaptive evolution".Proceedings of the American Philosophical Society.156(1): 45–57.JSTOR23558077.PMID23035389.
- ↑Kappelman J, Alçiçek MC, Kazanci N, Schultz M, Ozkul M, Sen S (Januarie 2008). "First Homo erectus from Turkey and implications for migrations into temperate Eurasia".American Journal of Physical Anthropology.135(1): 110–116.doi:10.1002/ajpa.20739.PMID18067194.
- ↑Best A, Kamilar JM (April 2018). "The evolution of eccrine sweat glands in human and nonhuman primates".Journal of Human Evolution.117:33–43.doi:10.1016/j.jhevol.2017.12.003.PMID29544622.
- ↑Pagel M, Bodmer W (2004). "The Evolution of Human Hairlessness: Cultural Adaptations and the Ectoparasite Hypothesis".Evolutionary Theory and Processes: Modern Horizons.Springer, Dordrecht. pp. 329–335.doi:10.1007/978-94-017-0443-4_17.ISBN978-94-017-0443-4.
- ↑Gile J (2010). "Naked Love: The Evolution of Human Hairlessness".Biological Theory.5(4): 326–336.doi:10.1162/BIOT_a_00062.S2CID84164968.
- ↑Coqueugniot H, Hublin JJ, Veillon F, Houët F, Jacob T (September 2004). "Early brain growth in Homo erectus and implications for cognitive ability".Nature.431(7006): 299–302.Bibcode:2004Natur.431..299C.doi:10.1038/nature02852.PMID15372030.S2CID4428043.
- ↑Steudel-Numbers KL (November 2006). "Energetics in Homo erectus and other early hominins: the consequences of increased lower-limb length".Journal of Human Evolution.51(5): 445–453.doi:10.1016/j.jhevol.2006.05.001.PMID16780923.
- ↑Kennedy GE (1985). "Bone thickness inHomo erectus".Journal of Human Evolution.14(8): 699–708.doi:10.1016/S0047-2484(85)80052-X.
- ↑Boaz N, Ciochon R (2004)."Headstrong Hominids".Natural History.113(1): 28–34.
- ↑61,061,1Willems EP, van Schaik CP (Augustus 2017). "The social organization of Homo ergaster: Inferences from anti-predator responses in extant primates".Journal of Human Evolution.109:11–21.doi:10.1016/j.jhevol.2017.05.003.PMID28688456.
- ↑Carotenuto F, Tsikaridze N, Rook L, Lordkipanidze D, Longo L, Condemi S, Raia P (Junie 2016). "Venturing out safely: The biogeography of Homo erectus dispersal out of Africa".Journal of Human Evolution.95:1–12.doi:10.1016/j.jhevol.2016.02.005.hdl:10356/82274.PMID27260171.
- ↑Simpson SW, Quade J, Levin NE, Butler R, Dupont-Nivet G, Everett M, Semaw S (November 2008). "A female Homo erectus pelvis from Gona, Ethiopia".Science.322(5904): 1089–1092.Bibcode:2008Sci...322.1089S.doi:10.1126/science.1163592.PMID19008443.S2CID22191315.
- ↑Plavcan JM (2012). "Body Size, Size Variation, and Sexual Size Dimorphism in EarlyHomo".Current Anthropology.53(S6): 309–423.doi:10.1086/667605.S2CID84095311.
- ↑Plavcan JM (2012). "Implications of Male and Female Contributions to Sexual Size Dimorphism for Inferring Behavior in the Hominin Fossil Record".International Journal of Primatology.33(6): 1364–1381.doi:10.1007/s10764-012-9642-z.S2CID17850676.
- ↑66,066,1Ben-Dor M, Gopher A, Hershkovitz I, Barkai R (2011)."Man the fat hunter: the demise of Homo erectus and the emergence of a new hominin lineage in the Middle Pleistocene (ca. 400 kyr) Levant".PLOS ONE.6(12): e28689.Bibcode:2011PLoSO...628689B.doi:10.1371/journal.pone.0028689.PMC3235142.PMID22174868.
- ↑Melamed Y, Kislev ME, Geffen E, Lev-Yadun S, Goren-Inbar N (Desember 2016)."The plant component of an Acheulian diet at Gesher Benot Ya'aqov, Israel".Proceedings of the National Academy of Sciences of the United States of America.113(51): 14674–14679.Bibcode:2016PNAS..11314674M.doi:10.1073/pnas.1607872113.PMC5187744.PMID27930293.
- ↑Steele TE (Junie 2010)."A unique hominin menu dated to 1.95 million years ago".Proceedings of the National Academy of Sciences of the United States of America.107(24): 10771–10772.Bibcode:2010PNAS..10710771S.doi:10.1073/pnas.1005992107.PMC2890732.PMID20534542.
- ↑Beck RB, Black L, Krieger LS, Naylor PC, Shabaka DI (1999).World History: Patterns of Interaction.Evanston, IL: McDougal Littell.ISBN978-0-395-87274-1.
- ↑Richards MP (Desember 2002)."A brief review of the archaeological evidence for Palaeolithic and Neolithic subsistence".European Journal of Clinical Nutrition.56(12): 1270–1278.doi:10.1038/sj.ejcn.1601646.PMID12494313.
- ↑de la Torre I (Julie 2016)."The origins of the Acheulean: past and present perspectives on a major transition in human evolution".Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences.371(1698): 20150245.doi:10.1098/rstb.2015.0245.PMC4920301.PMID27298475.
- ↑72,072,1Lepre CJ, Roche H, Kent DV, Harmand S, Quinn RL, Brugal JP, et al. (Augustus 2011). "An earlier origin for the Acheulian".Nature.477(7362): 82–85.Bibcode:2011Natur.477...82L.doi:10.1038/nature10372.PMID21886161.S2CID4419567.
- ↑Nowell A, Chang ML (2009)."The Case Against Sexual Selection as an Explanation of Handaxe Morphology"(PDF).PaleoAnthropology:77–88.
- ↑"Acheuleen" in dieWAT.Aanlyn byviva-afrikaans.org(intekening nodig). Besoek op 15 Mei 2023.
- ↑Choi K, Driwantoro D (2007). "Shell tool use by early members ofHomo erectusin Sangiran, central Java, Indonesia: cut mark evidence ".Journal of Archaeological Science.34(1): 48–58.Bibcode:2007JArSc..34...48C.doi:10.1016/j.jas.2006.03.013.
- ↑76,076,1Joordens JC, d'Errico F, Wesselingh FP, Munro S, de Vos J, Wallinga J, et al. (Februarie 2015). "Homo erectus at Trinil on Java used shells for tool production and engraving".Nature.518(7538): 228–231.Bibcode:2015Natur.518..228J.doi:10.1038/nature13962.PMID25470048.S2CID4461751.
- ↑Turner J (1996).History and Science of Knots.World Scientific. pp. 6–8.ISBN9789810224691.
- ↑James, Steven R. (Februarie 1989)."Hominid Use of Fire in the Lower and Middle Pleistocene: A Review of the Evidence"(PDF).Current Anthropology.University of Chicago Press.30(1): 1–26.doi:10.1086/203705.Geargiveer vanafdie oorspronklike(PDF)op 12 Desember 2015.Besoek op4 April2012.
- ↑Musée de Préhistoire Terra Amata."Le site acheuléen de Terra Amata"[The Acheulean site of Terra Amata].Musée de Préhistoire Terra Amata(in Frans).Besoek op10 Junie2022.
- ↑Leakey MD (1971).Olduvai Gorge: Volume 3, Excavations in Beds I and II, 1960-1963.Cambridge University Press. p. 24.ISBN9780521077231.
- ↑Ingold T (2000). "Building, dwelling, living: how animals and people make themselves at home in the world".The Perception of the Environment: Essays on Livelihood, Dwelling and Skill.Psychology Press. p. 184.ISBN9780415228329.
- ↑Sklenář K (1987). "The Lower Paleolithic Dwelling Structure at Přezletice and its Significance".Anthropologie.25(2): 101–103.JSTOR26294864.
- ↑Ullman M, Hovers E, Goren-Inbar N, Frumkin A (2013)."Levantine cave dwellers: geographic and environmental aspects of early humans use of caves, case study from Wadi Amud, northern Israel".International Congress of Speleology.1.
- ↑Toups MA, Kitchen A, Light JE, Reed DL (Januarie 2011)."Origin of clothing lice indicates early clothing use by anatomically modern humans in Africa".Molecular Biology and Evolution.28(1): 29–32.doi:10.1093/molbev/msq234.PMC3002236.PMID20823373.
- ↑"New evidence in search for the mysterious Denisovans".ScienceDaily(in Engels). 23 Maart 2021.Besoek op30 Maart2021.
- ↑86,086,1Bednarik RG (1999). "Pleistocene seafaring in the Mediterranean".Anthropologie.37(3): 275–282.JSTOR26294895.
- ↑Bednarik RG (1998)."An experiment in Pleistocene seafaring"(PDF).The International Journal of Nautical Archaeology.27(2): 139–149.doi:10.1111/j.1095-9270.1998.tb00797.x.
- ↑Détroit F, Mijares AS, Corny J, Daver G, Zanolli C, Dizon E, et al. (April 2019)."A new species of Homo from the Late Pleistocene of the Philippines"(PDF).Nature.568(7751): 181–186.Bibcode:2019Natur.568..181D.doi:10.1038/s41586-019-1067-9.PMID30971845.S2CID106411053.
- ↑Spikins P, Needham A, Wright B, Dytham C, Gatta M, Hitchens G (2019)."Living to fight another day: The ecological and evolutionary significance of Neanderthal healthcare".Quaternary Science Reviews.217:98–118.Bibcode:2019QSRv..217...98S.doi:10.1016/j.quascirev.2018.08.011.
- ↑Lordkipanidze D, Vekua A, Ferring R, Rightmire GP, Agusti J, Kiladze G, et al. (April 2005). "Anthropology: the earliest toothless hominin skull".Nature.434(7034): 717–718.Bibcode:2005Natur.434..717L.doi:10.1038/434717b.PMID15815618.S2CID52800194.
- ↑Haeusler M, Schiess R, Boeni T (Februarie 2013)."Evidence for juvenile disc herniation in a homo erectus boy skeleton"(PDF).Spine.38(3): E123–E128.doi:10.1097/BRS.0b013e31827cd245.PMID23154836.S2CID11534863.
- ↑Henshilwood CS, d'Errico F, Watts I (Julie 2009). "Engraved ochres from the Middle Stone Age levels at Blombos Cave, South Africa".Journal of Human Evolution.57(1): 27–47.doi:10.1016/j.jhevol.2009.01.005.PMID19487016.
- ↑93,093,193,2Dickson DB (1992).The Dawn of Belief: Religion in the Upper Paleolithic of Southwestern Europe.University of Arizona Press. pp. 40–46.ISBN978-0-8165-1336-9.
- ↑94,094,1Morriss-Kay GM (Februarie 2010)."The evolution of human artistic creativity".Journal of Anatomy.216(2): 158–176.doi:10.1111/j.1469-7580.2009.01160.x.PMC2815939.PMID19900185.
- ↑d'Errico F, Nowell A (2000). "A New Look at the Berekhat Ram Figurine: Implications for the Origins of Symbolism".Cambridge Archaeological Journal.10(1): 123–167.doi:10.1017/S0959774300000056.S2CID163138037.
- ↑96,096,1Watts I (2014). "The red thread: pigment use and the evolution of collective ritual".The Social Origins of Language.Oxford University Press. pp. 222–223.ISBN978-0-19-966533-4.
- ↑de Lumley H, Boone Y (1976). "Les structures d'habitat au Paléolithique moyen" [Housing structures from the lower Paleolithic]. In de Lumley H, Guilaine J (reds.).La Préhistoire française: Les civilisations paléolithiques et mésolithiques de la France[French prehistory: the Paleolithic and Mesolithic civilizations of France]. Éditions du Centre national de la recherche scientifique.ISBN978-2-222-01968-8.
- ↑Wreschner EE, Bolton R, Butzer KW, Delporte H, Häusler A, Heinrich A, et al. (1980)."Red Ochre and Human Evolution: A Case for Discussion"(PDF).Current Anthropology.21(5): 632–633.doi:10.1086/202541.JSTOR2741829.S2CID88099778.
- ↑Latimer B, Ohman J (2001). "Axial dysplasia inHomo erectus".Journal of Human Evolution.40.
- ↑(2006) "Language and empathy in Homo erectus: Behaviors suggested by a modern spinal cord from Dmanisi, but not Nariokotome."inAnnual meeting of the Paleoanthroplogy Society..
- ↑Luef EM (2018). "Tracing the human brain's classical language areas in extant and extinct hominids".The talking species: Perspectives on the evolutionary, neuronal and cultural foundations of language.Uni-Press Graz.ISBN978-3-902666-52-9.
- ↑Capasso L, Michetti E, D'Anastasio R (Desember 2008). "A Homo erectus hyoid bone: possible implications for the origin of the human capability for speech".Collegium Antropologicum.32(4): 1007–1011.PMID19149203.
- ↑Padmanaban D (6 November 2020)."Climate Change May Have Been a Major Driver of Ancient Hominin Extinctions".SAPIENS.Besoek op9 November2020.
- ↑"Climate change likely drove early human species to extinction, modeling study suggests".phys.org(in Engels).Besoek op9 November2020.
- ↑Raia P, Mondanaro A, Melchionna M, Di Febbraro M, Diniz-Filho JA, Rangel TF, et al. (23 Oktober 2020)."Past Extinctions of Homo Species Coincided with Increased Vulnerability to Climatic Change".One Earth(in Engels).3(4): 480–490.Bibcode:2020OEart...3..480R.doi:10.1016/j.oneear.2020.09.007.hdl:2158/1211341.ISSN2590-3330.
- ↑Ni, Xijun; Ji, Qiang; Wu, Wensheng; Shao, Qingfeng; Ji, Yannan; Zhang, Chi; Liang, Lei; Ge, Junyi; Guo, Zhen; Li, Jinhua; Li, Qiang; Grün, Rainer; Stringer, Chris (28 Augustus 2021)."Massive cranium from Harbin in northeastern China establishes a new Middle Pleistocene human lineage".The Innovation(in English).2(3): 100130.Bibcode:2021Innov...200130N.doi:10.1016/j.xinn.2021.100130.ISSN2666-6758.PMC8454562.
{{cite journal}}:AS1-onderhoud: onerkende taal (link)
Nog leesstof
[wysig|wysig bron]- Leakey R, Walker A (November 1985). "Homo Erectus Unearthed".National Geographic.Vol. 168, no. 5. pp. 624–629.ISSN0027-9358.OCLC643483454.
- Sigmon BA, Cybulski JS (1981).Homo erectus: Papers in Honor of Davidson Black.University of Toronto Press.JSTOR10.3138/j.ctvcj2jdw.11.
- Theunissen B, Theunissen LT (2012).Eugène Dubois and the Ape-Man from Java.Springer Netherlands.ISBN9789400922099.
Eksterne skakels
[wysig|wysig bron]- Archaeology Info
- Homo erectus– The Smithsonian Institution
 Wikimedia Commonshet meer media in die kategorieHomo erectus.
Wikimedia Commonshet meer media in die kategorieHomo erectus.
 Wikispecieshet meer inligting verwant aanHomo erectus
Wikispecieshet meer inligting verwant aanHomo erectus Hierdie artikel is vertaal uit dieEngelse Wikipedia
Hierdie artikel is vertaal uit dieEngelse Wikipedia