Convergent evolutionis the independentevolutionof similar features in species of different periods or epochs in time. Convergent evolution createsanalogous structuresthat have similar form or function but were not present in thelast common ancestorof those groups. Thecladisticterm for the same phenomenon ishomoplasy.Therecurrent evolutionof flight is a classic example, as flyinginsects,birds,pterosaurs,andbatshave independently evolved the useful capacity of flight. Functionally similar features that have arisen through convergent evolution areanalogous,whereashomologousstructures or traits have a common origin but can have dissimilar functions. Bird, bat, and pterosaurwingsare analogous structures, but their forelimbs are homologous, sharing an ancestral state despite serving different functions.


The opposite of convergence isdivergent evolution,where related species evolve different traits. Convergent evolution is similar toparallel evolution,which occurs when two independent species evolve in the same direction and thus independently acquire similar characteristics; for instance,gliding frogshave evolved in parallel from multiple types oftree frog.
Many instances of convergent evolution are known inplants,including the repeated development ofC4photosynthesis,seed dispersalby fleshyfruitsadapted to be eaten by animals, andcarnivory.
Overview
edit
In morphology, analogous traits arise when different species live in similar ways and/or a similar environment, and so face the same environmental factors. When occupying similarecological niches(that is, a distinctive way of life) similar problems can lead to similar solutions.[1][2][3]The British anatomistRichard Owenwas the first to identify the fundamental difference between analogies andhomologies.[4]
In biochemistry, physical and chemical constraints onmechanismshave caused someactive sitearrangements such as thecatalytic triadto evolve independently in separateenzyme superfamilies.[5]
In his 1989 bookWonderful Life,Stephen Jay Gouldargued that if one could "rewind the tape of life [and] the same conditions were encountered again, evolution could take a very different course."[6]Simon Conway Morrisdisputes this conclusion, arguing that convergence is a dominant force in evolution, and given that the same environmental and physical constraints are at work, life will inevitably evolve toward an "optimum" body plan, and at some point, evolution is bound to stumble upon intelligence, a trait presently identified with at leastprimates,corvids,andcetaceans.[7]
Distinctions
editCladistics
editIn cladistics, a homoplasy is a trait shared by two or moretaxafor any reason other than that they share a common ancestry. Taxa which do share ancestry are part of the sameclade;cladistics seeks to arrange them according to their degree of relatedness to describe theirphylogeny.Homoplastic traits caused by convergence are therefore, from the point of view of cladistics, confounding factors which could lead to an incorrect analysis.[8][9][10][11]
Atavism
editIn some cases, it is difficult to tell whether a trait has been lost and then re-evolved convergently, or whether a gene has simply been switched off and then re-enabled later. Such a re-emerged trait is called anatavism.From a mathematical standpoint, an unused gene (selectively neutral) has a steadily decreasingprobabilityof retaining potential functionality over time. The time scale of this process varies greatly in different phylogenies; in mammals and birds, there is a reasonable probability of remaining in the genome in a potentially functional state for around 6 million years.[12]
Parallel vs. convergent evolution
edit
When two species are similar in a particular character, evolution is defined as parallel if the ancestors were also similar, and convergent if they were not.[b]Some scientists have argued that there is a continuum between parallel and convergent evolution,[13][14][15][16]while others maintain that despite some overlap, there are still important distinctions between the two.[17][18]
When the ancestral forms are unspecified or unknown, or the range of traits considered is not clearly specified, the distinction between parallel and convergent evolution becomes more subjective. For instance, the striking example of similar placental and marsupial forms is described byRichard DawkinsinThe Blind Watchmakeras a case of convergent evolution, because mammals on each continent had a long evolutionary history prior to the extinction of the dinosaurs under which to accumulate relevant differences.[19]
At molecular level
edit
Proteins
editProtease active sites
editTheenzymologyofproteasesprovides some of the clearest examples of convergent evolution. These examples reflect the intrinsic chemical constraints on enzymes, leading evolution to converge on equivalent solutions independently and repeatedly.[5][20]
Serine and cysteine proteases use different amino acid functional groups (alcohol or thiol) as anucleophile.In order to activate that nucleophile, they orient an acidic and a basic residue in acatalytic triad.The chemical and physical constraints onenzyme catalysishave caused identical triad arrangements to evolve independently more than 20 times in differentenzyme superfamilies.[5]
Threonine proteasesuse the amino acid threonine as their catalyticnucleophile.Unlike cysteine and serine, threonine is asecondary alcohol(i.e. has a methyl group). The methyl group of threonine greatly restricts the possible orientations of triad and substrate, as the methyl clashes with either the enzyme backbone or the histidine base. Consequently, most threonine proteases use an N-terminal threonine in order to avoid suchsteric clashes. Several evolutionarily independentenzyme superfamilieswith differentprotein foldsuse the N-terminal residue as a nucleophile. This commonality ofactive sitebut difference of protein fold indicates that the active site evolved convergently in those families.[5][21]
Cone snail and fish insulin
editConus geographusproduces a distinct form ofinsulinthat is more similar to fish insulin protein sequences than to insulin from more closely related molluscs, suggesting convergent evolution,[22]though with the possibility ofhorizontal gene transfer.[23]
Ferrous iron uptake via protein transporters in land plants and chlorophytes
editDistant homologues of the metal ion transportersZIPinland plantsandchlorophyteshave converged in structure, likely to take up Fe2+efficiently. The IRT1 proteins fromArabidopsis thalianaandricehave extremely different amino acid sequences fromChlamydomonas's IRT1, but their three-dimensional structures are similar, suggesting convergent evolution.[24]
Na+,K+-ATPase and Insect resistance to cardiotonic steroids
editMany examples of convergent evolution exist in insects in terms of developing resistance at a molecular level to toxins. One well-characterized example is the evolution of resistance to cardiotonic steroids (CTSs) via amino acid substitutions at well-defined positions of the α-subunit ofNa+,K+-ATPase(ATPalpha). Variation in ATPalpha has been surveyed in various CTS-adapted species spanning six insect orders.[25][26][27]Among 21 CTS-adapted species, 58 (76%) of 76 amino acid substitutions at sites implicated in CTS resistance occur in parallel in at least two lineages.[27]30 of these substitutions (40%) occur at just two sites in the protein (positions 111 and 122). CTS-adapted species have also recurrently evolvedneo-functionalizedduplications of ATPalpha, with convergent tissue-specific expression patterns.[25][27]
Nucleic acids
editConvergence occurs at the level ofDNAand theamino acidsequences produced bytranslatingstructural genesintoproteins.Studies have found convergence in amino acid sequences in echolocating bats and the dolphin;[28]among marine mammals;[29]between giant and red pandas;[30]and between the thylacine and canids.[31]Convergence has also been detected in a type ofnon-coding DNA,cis-regulatory elements,such as in their rates of evolution; this could indicate eitherpositive selectionor relaxedpurifying selection.[32][33]
In animal morphology
edit
Bodyplans
editSwimming animals includingfishsuch asherrings,marine mammalssuch asdolphins,andichthyosaurs(of the Mesozoic) all converged on the same streamlined shape.[34][35]A similar shape and swimming adaptations are even present in molluscs, such asPhylliroe.[36]The fusiform bodyshape (a tube tapered at both ends) adopted by many aquatic animals is an adaptation to enable them totravel at high speedin a highdragenvironment.[37]Similar body shapes are found in theearless sealsand theeared seals:they still have four legs, but these are strongly modified for swimming.[38]
The marsupial fauna of Australia and the placental mammals of the Old World have several strikingly similar forms, developed in two clades, isolated from each other.[7]The body, and especially the skull shape, of thethylacine(Tasmanian tiger or Tasmanian wolf) converged with those ofCanidaesuch as the red fox,Vulpes vulpes.[39]
- Convergence ofmarsupialandplacentalmammals
-
 Red foxskeleton
Red foxskeleton -
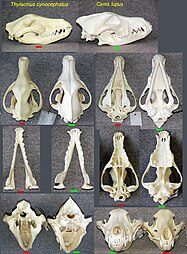 Skulls ofthylacine(left),timber wolf(right)
Skulls ofthylacine(left),timber wolf(right) -
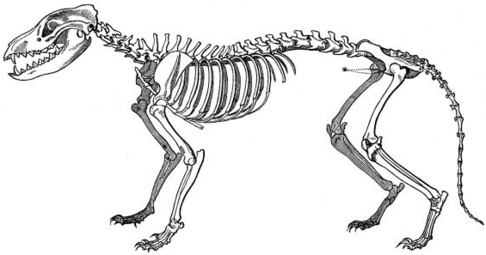 Thylacineskeleton
Thylacineskeleton



Echolocation
editAs a sensory adaptation,echolocationhas evolved separately incetaceans(dolphins and whales) and bats, but from the same genetic mutations.[40]
Electric fishes
editTheGymnotiformesof South America and theMormyridaeof Africa independently evolvedpassive electroreception(around 119 and 110 million years ago, respectively). Around 20 million years after acquiring that ability, both groups evolved activeelectrogenesis,producing weak electric fields to help them detect prey.[41]
- Convergence ofweakly electric fishes
-
 Gymnotiform electrolocation waveform
Gymnotiform electrolocation waveform -
 Agymnotiformelectric fish of South America
Agymnotiformelectric fish of South America -
 Amormyridelectric fish of Africa
Amormyridelectric fish of Africa -
Mormyrid electrolocation waveform


Eyes
edit
One of the best-known examples of convergent evolution is the camera eye ofcephalopods(such as squid and octopus),vertebrates(including mammals) andcnidaria(such as jellyfish).[43]Their last common ancestor had at most a simple photoreceptive spot, but a range of processes led to theprogressive refinement of camera eyes—with one sharp difference: the cephalopod eye is "wired" in the opposite direction, with blood and nerve vessels entering from the back of the retina, rather than the front as in vertebrates. As a result, vertebrates have ablind spot.[7]
Flight
edit
Birdsandbatshavehomologouslimbs because they are both ultimately derived from terrestrialtetrapods,but their flight mechanisms are only analogous, so their wings are examples of functional convergence. The two groups have independently evolved their own means of powered flight. Their wings differ substantially in construction. The bat wing is a membrane stretched across four extremely elongated fingers and the legs. The airfoil of the bird wing is made offeathers,strongly attached to the forearm (the ulna) and the highly fused bones of the wrist and hand (thecarpometacarpus), with only tiny remnants of two fingers remaining, each anchoring a single feather. So, while the wings of bats and birds are functionally convergent, they are not anatomically convergent.[3][44]Birds and bats also share a high concentration ofcerebrosidesin the skin of their wings. This improves skin flexibility, a trait useful for flying animals; other mammals have a far lower concentration.[45]The extinctpterosaursindependently evolved wings from their fore- and hindlimbs, whileinsectshavewingsthat evolved separately from different organs.[46]
Flying squirrelsandsugar glidersare much alike in their body plans, with gliding wings stretched between their limbs, but flying squirrels are placental mammals while sugar gliders are marsupials, widely separated within the mammal lineage from the placentals.[47]
Hummingbird hawk-mothsandhummingbirdshave evolved similar flight and feeding patterns.[48]
Insect mouthparts
editInsect mouthparts show many examples of convergent evolution. The mouthparts of different insect groups consist of a set ofhomologousorgans, specialised for the dietary intake of that insect group. Convergent evolution of many groups of insects led from original biting-chewing mouthparts to different, more specialised, derived function types. These include, for example, theproboscisof flower-visiting insects such asbeesandflower beetles,[49][50][51]or the biting-sucking mouthparts of blood-sucking insects such asfleasandmosquitos.
Opposable thumbs
editOpposable thumbsallowing the grasping of objects are most often associated withprimates,like humans and other apes, monkeys, and lemurs. Opposable thumbs also evolved ingiant pandas,but these are completely different in structure, having six fingers including the thumb, which develops from a wrist bone entirely separately from other fingers.[52]
Primates
edit Despite the similar lightening ofskin colourafter movingout of Africa,different genes were involved in European (left) and East Asian (right) lineages. Despite the similar lightening ofskin colourafter movingout of Africa,different genes were involved in European (left) and East Asian (right) lineages.
| ||



Convergent evolution in humans includes blue eye colour and light skin colour.[53]When humans migratedout of Africa,they moved to more northern latitudes with less intense sunlight.[53]It was beneficial to them to reduce theirskin pigmentation.[53]It appears certain that there was some lightening of skin colourbeforeEuropean and East Asian lineages diverged, as there are some skin-lightening genetic differences that are common to both groups.[53]However, after the lineages diverged and became genetically isolated, the skin of both groups lightened more, and that additional lightening was due todifferentgenetic changes.[53]
| Humans | Lemurs | ||
|---|---|---|---|
| Despite the similarity of appearance, the genetic basis of blue eyes is different in humans andlemurs. | |||




Lemursandhumansare both primates. Ancestral primates had brown eyes, as most primates do today. The genetic basis of blue eyes in humans has been studied in detail and much is known about it. It is not the case that onegene locusis responsible, say with brown dominant to blueeye colour.However, a single locus is responsible for about 80% of the variation. In lemurs, the differences between blue and brown eyes are not completely known, but the same gene locus is not involved.[54]
In plants
edit
The annual life-cycle
editWhile most plant species areperennial,about 6% follow anannuallife cycle, living for only one growing season.[55]The annual life cycle independently emerged in over 120 plant families of angiosperms.[56][57]The prevalence of annual species increases under hot-dry summer conditions in the four species-rich families of annuals (Asteraceae,Brassicaceae,Fabaceae,andPoaceae), indicating that the annual life cycle is adaptive.[55][58]
Carbon fixation
editC4photosynthesis,one of the three major carbon-fixing biochemical processes, hasarisen independently up to 40 times.[59][60]About 7,600 plant species ofangiospermsuse C4carbon fixation, with manymonocotsincluding 46% of grasses such asmaizeandsugar cane,[61][62]anddicotsincluding several species in theChenopodiaceaeand theAmaranthaceae.[63][64]
Fruits
editFruitswith a wide variety of structural origins have converged to become edible.Applesarepomeswith fivecarpels;their accessory tissues form the apple's core, surrounded by structures from outside the botanical fruit, thereceptacleorhypanthium.Other edible fruits include other plant tissues;[65]the fleshy part of atomatois the walls of thepericarp.[66]This implies convergent evolution under selective pressure, in this case the competition forseed dispersalby animals through consumption of fleshy fruits.[67]
Seed dispersal by ants (myrmecochory) has evolved independently more than 100 times, and is present in more than 11,000 plant species. It is one of the most dramatic examples of convergent evolution in biology.[68]
Carnivory
edit
Carnivoryhas evolved multiple times independently in plants in widely separated groups. In three species studied,Cephalotus follicularis,Nepenthes alataandSarracenia purpurea,there has been convergence at the molecular level. Carnivorous plants secreteenzymesinto the digestive fluid they produce. By studyingphosphatase,glycoside hydrolase,glucanase,RNAseandchitinaseenzymesas well as apathogenesis-related proteinand athaumatin-related protein, the authors found many convergentamino acidsubstitutions. These changes were not at the enzymes' catalytic sites, but rather on the exposed surfaces of the proteins, where they might interact with other components of the cell or the digestive fluid. The authors also found thathomologous genesin the non-carnivorous plantArabidopsis thalianatend to have their expression increased when the plant is stressed, leading the authors to suggest that stress-responsive proteins have often been co-opted[c]in the repeated evolution of carnivory.[69]
Methods of inference
edit
Phylogenetic reconstruction andancestral state reconstructionproceed by assuming that evolution has occurred without convergence. Convergent patterns may, however, appear at higher levels in a phylogenetic reconstruction, and are sometimes explicitly sought by investigators. The methods applied to infer convergent evolution depend on whether pattern-based or process-based convergence is expected. Pattern-based convergence is the broader term, for when two or more lineages independently evolve patterns of similar traits. Process-based convergence is when the convergence is due to similar forces ofnatural selection.[70]
Pattern-based measures
editEarlier methods for measuring convergence incorporate ratios of phenotypic andphylogeneticdistance by simulating evolution with aBrownian motionmodel of trait evolution along a phylogeny.[71][72]More recent methods also quantify the strength of convergence.[73]One drawback to keep in mind is that these methods can confuse long-term stasis with convergence due to phenotypic similarities. Stasis occurs when there is little evolutionary change among taxa.[70]
Distance-based measures assess the degree of similarity between lineages over time. Frequency-based measures assess the number of lineages that have evolved in a particular trait space.[70]
Process-based measures
editMethods to infer process-based convergence fit models of selection to a phylogeny and continuous trait data to determine whether the same selective forces have acted upon lineages. This uses theOrnstein–Uhlenbeck processto test different scenarios of selection. Other methods rely on ana priorispecification of where shifts in selection have occurred.[74]
See also
edit- Incomplete lineage sorting– Characteristic of phylogenetic analysis: the presence of multiple alleles in ancestral populations might lead to the impression that convergent evolution has occurred.
- Carcinisation– Evolution of crustaceans into crab-like forms
- Morphology (biology)– Study of external forms and structures of organisms
- Iterative evolution– The repeated evolution of a specific trait or body plan from the same ancestral lineage at different points in time.
- Elvis taxon– Misidentification of later taxon superficially resembling earlier extinct taxon
- Breeding back– A form of selective breeding to recreate the traits of an extinct species, but the genome will differ from the original species.
- Orthogenesis(contrastable with convergent evolution; involves teleology)
- Contingency (evolutionary biology)– effect of evolutionary history on outcomes
Notes
edit- ^However,evolutionary developmental biologyhas identifieddeep homologybetween insect and mammal body plans, to the surprise of many biologists.
- ^However, all organisms share a common ancestor more or less recently, so the question of how far back to look in evolutionary time and how similar the ancestors need to be for one to consider parallel evolution to have taken place is not entirely resolved within evolutionary biology.
- ^The prior existence of suitable structures has been calledpre-adaptationorexaptation.
References
edit- ^Kirk, John Thomas Osmond (2007).Science & Certainty.Csiro Publishing. p. 79.ISBN978-0-643-09391-1.Archivedfrom the original on 15 February 2017.Retrieved23 January2017.
evolutionary convergence, which, quoting.. Simon Conway Morris.. is the 'recurring tendency of biological organization to arrive at the same "solution" to a particular "need"... the 'Tasmanian tiger'.. looked and behaved like a wolf and occupied a similar ecological niche, but was in fact a marsupial not a placental mammal.
- ^Reece, J.; Meyers, N.; Urry, L.; Cain, M.; Wasserman, S.; Minorsky, P.; Jackson, R.; Cooke, B. (5 September 2011).Cambell Biology, 9th Edition.Pearson. p. 586.ISBN978-1-4425-3176-5.
- ^ab"Homologies and analogies".University of California Berkeley.Archivedfrom the original on 19 November 2016.Retrieved10 January2017.
- ^Thunstad, Erik (2009).Darwins teori, evolusjon gjennom 400 år(in Norwegian). Oslo, Norway: Humanist forlag. p. 404.ISBN978-82-92622-53-7.
- ^abcdBuller, A. R.; Townsend, C. A. (19 February 2013)."Intrinsic evolutionary constraints on protease structure, enzyme acylation, and the identity of the catalytic triad".Proceedings of the National Academy of Sciences of the United States of America.110(8): E653–61.Bibcode:2013PNAS..110E.653B.doi:10.1073/pnas.1221050110.PMC3581919.PMID23382230.
- ^Gould, S.J.(1989).Wonderful Life: The Burgess Shale and the Nature of History.W.W. Norton. pp.282–285.ISBN978-0-09-174271-3.
- ^abcConway Morris, Simon(2005).Life's solution: inevitable humans in a lonely universe.Cambridge University Press. pp.164, 167, 170 and 235.ISBN978-0-521-60325-6.OCLC156902715.
- ^Chirat, R.; Moulton, D. E.; Goriely, A. (2013)."Mechanical basis of morphogenesis and convergent evolution of spiny seashells".Proceedings of the National Academy of Sciences.110(15): 6015–6020.Bibcode:2013PNAS..110.6015C.doi:10.1073/pnas.1220443110.PMC3625336.PMID23530223.
- ^Lomolino, M; Riddle, B; Whittaker, R; Brown, J (2010).Biogeography, Fourth Edition.Sinauer Associates. p. 426.ISBN978-0-87893-494-2.
- ^West-Eberhard, Mary Jane(2003).Developmental Plasticity and Evolution.Oxford University Press. pp. 353–376.ISBN978-0-19-512235-0.
- ^Sanderson, Michael J.; Hufford, Larry (1996).Homoplasy: The Recurrence of Similarity in Evolution.Academic Press. pp. 330, and passim.ISBN978-0-08-053411-4.Archivedfrom the original on 14 February 2017.Retrieved21 January2017.
- ^Collin, R.; Cipriani, R. (2003)."Dollo's law and the re-evolution of shell coiling".Proceedings of the Royal Society B.270(1533): 2551–2555.doi:10.1098/rspb.2003.2517.PMC1691546.PMID14728776.
- ^Arendt, J; Reznick, D (January 2008). "Convergence and parallelism reconsidered: what have we learned about the genetics of adaptation?".Trends in Ecology & Evolution.23(1): 26–32.Bibcode:2008TEcoE..23...26A.doi:10.1016/j.tree.2007.09.011.PMID18022278.
- ^Waters, Jonathan M.; McCulloch, Graham A. (2021). "Reinventing the wheel? Reassessing the roles of gene flow, sorting and convergence in repeated evolution".Molecular Ecology.30(17): 4162–4172.Bibcode:2021MolEc..30.4162W.doi:10.1111/mec.16018.ISSN1365-294X.PMID34133810.S2CID235460165.
- ^Cerca, José (October 2023). "Understanding natural selection and similarity: Convergent, parallel and repeated evolution".Molecular Ecology.32(20): 5451–5462.Bibcode:2023MolEc..32.5451C.doi:10.1111/mec.17132.PMID37724599.
- ^Bohutínská, Magdalena; Peichel, Catherine L. (April 2024). "Divergence time shapes gene reuse during repeated adaptation".Trends in Ecology & Evolution.39(4): 396–407.Bibcode:2024TEcoE..39..396B.doi:10.1016/j.tree.2023.11.007.PMID38155043.
- ^Pearce, T. (10 November 2011)."Convergence and Parallelism in Evolution: A Neo-Gouldian Account".The British Journal for the Philosophy of Science.63(2): 429–448.doi:10.1093/bjps/axr046.
- ^Zhang, J.; Kumar, S. (1997)."Detection of convergent and parallel evolution at the amino acid sequence level".Mol. Biol. Evol.14(5): 527–36.doi:10.1093/oxfordjournals.molbev.a025789.PMID9159930.
- ^Dawkins, Richard(1986).The Blind Watchmaker.W. W. Norton. pp.100–106.ISBN978-0-393-31570-7.
- ^Dodson, G.; Wlodawer, A. (September 1998). "Catalytic triads and their relatives".Trends in Biochemical Sciences.23(9): 347–52.doi:10.1016/S0968-0004(98)01254-7.PMID9787641.
- ^Ekici, O. D.; Paetzel, M.; Dalbey, R. E. (December 2008)."Unconventional serine proteases: variations on the catalytic Ser/His/Asp triad configuration".Protein Science.17(12): 2023–37.doi:10.1110/ps.035436.108.PMC2590910.PMID18824507.
- ^Safavi-Hemami, Helena; Gajewiak, Joanna; Karanth, Santhosh; Robinson, Samuel D.; Ueberheide, Beatrix; Douglass, Adam D.; Schlegel, Amnon; Imperial, Julita S.; Watkins, Maren; Bandyopadhyay, Pradip K.; Yandell, Mark; Li, Qing; Purcell, Anthony W.; Norton, Raymond S.; Ellgaard, Lars; Olivera, Baldomero M. (10 February 2015)."Specialized insulin is used for chemical warfare by fish-hunting cone snails".Proceedings of the National Academy of Sciences.112(6): 1743–1748.Bibcode:2015PNAS..112.1743S.doi:10.1073/pnas.1423857112.PMC4330763.PMID25605914.
- ^Martin, J. P.; Fridovich, I (1981)."Evidence for a natural gene-transfer from the ponyfish to its bioluminescent bacterial symbiontPhotobacter leiognathi— the close relationship between bacteriocuprein and the copper-zinc superoxide-dismutase of teleost fishes ".J. Biol. Chem.256(12): 6080–6089.doi:10.1016/S0021-9258(19)69131-3.PMID6787049.
- ^Rodrigues, Wenderson Felipe Costa; Lisboa, Ayrton Breno P.; Lima, Joni Esrom; Ricachenevsky, Felipe Klein; Del-Bem, Luiz-Eduardo (10 January 2023)."Ferrous iron uptake via IRT1 / ZIP evolved at least twice in green plants".New Phytologist.237(6): 1951–1961.doi:10.1111/nph.18661.PMID36626937.
- ^abZhen, Ying; Aardema, Matthew L.; Medina, Edgar M.; Schumer, Molly; Andolfatto, Peter (28 September 2012)."Parallel Molecular Evolution in an Herbivore Community".Science.337(6102): 1634–1637.Bibcode:2012Sci...337.1634Z.doi:10.1126/science.1226630.ISSN0036-8075.PMC3770729.PMID23019645.
- ^Dobler, S., Dalla, S., Wagschal, V., & Agrawal, A. A. (2012). Community-wide convergent evolution in insect adaptation to toxic cardenolides by substitutions in the Na,K-ATPase. Proceedings of the National Academy of Sciences, 109(32), 13040–13045.https://doi.org/10.1073/pnas.1202111109
- ^abcYang, L; Ravikanthachari, N; Mariño-Pérez, R; Deshmukh, R; Wu, M; Rosenstein, A; Kunte, K; Song, H; Andolfatto, P. (2019)."Predictability in the evolution of Orthopteran cardenolide insensitivity".Philosophical Transactions of the Royal Society of London, Series B.374(1777): 20180246.doi:10.1098/rstb.2018.0246.PMC6560278.PMID31154978.
- ^Liu, Zhen; Qi, Fei-Yan; Zhou, Xin; Ren, Hai-Qing; Shi, Peng (2014)."Parallel Sites Implicate Functional Convergence of the Hearing Gene Prestin among Echolocating Mammals".Molecular Biology and Evolution.31(9): 2415–2424.doi:10.1093/molbev/msu194.ISSN1537-1719.PMID24951728.
- ^Foote, Andrew D.; Liu, Yue; Thomas, Gregg W. C.; Vinař, Tomáš; Alföldi, Jessica; Deng, Jixin; Dugan, Shannon; Elk, Cornelis E. van; Hunter, Margaret E. (March 2015)."Convergent evolution of the genomes of marine mammals".Nature Genetics.47(3): 272–275.doi:10.1038/ng.3198.PMC4644735.PMID25621460.
- ^Hu, Yibo; Wu, Qi; Ma, Shuai; Ma, Tianxiao; Shan, Lei; Wang, Xiao; Nie, Yonggang; Ning, Zemin; Yan, Li (January 2017)."Comparative genomics reveals convergent evolution between the bamboo-eating giant and red pandas".Proceedings of the National Academy of Sciences of the United States of America.114(5): 1081–1086.Bibcode:2017PNAS..114.1081H.doi:10.1073/pnas.1613870114.PMC5293045.PMID28096377.
- ^Feigin, Charles Y.; Newton, Axel H.; Doronina, Liliya; Schmitz, Jürgen; Hipsley, Christy A.; Mitchell, Kieren J.; Gower, Graham; Llamas, Bastien; Soubrier, Julien (January 2018)."Genome of the Tasmanian tiger provides insights into the evolution and demography of an extinct marsupial carnivore".Nature Ecology & Evolution.2(1): 182–192.doi:10.1038/s41559-017-0417-y.PMID29230027.
- ^Partha, Raghavendran; Chauhan, Bharesh K; Ferreira, Zelia; Robinson, Joseph D; Lathrop, Kira; Nischal, Ken K.; Chikina, Maria; Clark, Nathan L. (October 2017)."Subterranean mammals show convergent regression in ocular genes and enhancers, along with adaptation to tunneling".eLife.6.doi:10.7554/eLife.25884.PMC5643096.PMID29035697.
- ^Sackton, TB; Grayson, P; Cloutier, A; Hu, Z; Liu, JS; Wheeler, NE; Gardner, PP; Clarke, JA; Baker, AJ; Clamp, M; Edwards, SV (5 April 2019)."Convergent regulatory evolution and loss of flight in paleognathous birds".Science.364(6435): 74–78.Bibcode:2019Sci...364...74S.doi:10.1126/science.aat7244.PMID30948549.S2CID96435050.
- ^"How do analogies evolve?".University of California Berkeley.Archivedfrom the original on 2 April 2017.Retrieved26 January2017.
- ^Selden, Paul; Nudds, John (2012).Evolution of Fossil Ecosystems(2nd ed.). CRC Press. p. 133.ISBN978-1-84076-623-3.Archivedfrom the original on 15 February 2017.Retrieved26 January2017.
- ^Helm, R. R. (18 November 2015)."Meet Phylliroe: the sea slug that looks and swims like a fish".Deep Sea News.Archivedfrom the original on 26 July 2019.Retrieved26 July2019.
- ^Ballance, Lisa (2016)."The Marine Environment as a Selective Force for Secondary Marine Forms"(PDF).UCSD.Archived(PDF)from the original on 2 February 2017.Retrieved19 September2019.
- ^Lento, G. M.; Hickson, R. E.; Chambers, G. K.; Penny, D. (1995)."Use of spectral analysis to test hypotheses on the origin of pinnipeds".Molecular Biology and Evolution.12(1): 28–52.doi:10.1093/oxfordjournals.molbev.a040189.PMID7877495.
- ^Werdelin, L. (1986). "Comparison of Skull Shape in Marsupial and Placental Carnivores".Australian Journal of Zoology.34(2): 109–117.doi:10.1071/ZO9860109.
- ^Liu, Yang; Cotton, James A.; Shen, Bin; Han, Xiuqun; Rossiter, Stephen J.; Zhang, Shuyi (1 January 2010)."Convergent sequence evolution between echolocating bats and dolphins".Current Biology.20(2): R53–R54.Bibcode:2010CBio...20..R53L.doi:10.1016/j.cub.2009.11.058.PMID20129036.S2CID16117978.
- ^Lavoué, Sébastien; Miya, Masaki; Arnegard, Matthew E.; Sullivan, John P.; Hopkins, Carl D.; Nishida, Mutsumi (14 May 2012)."Comparable Ages for the Independent Origins of Electrogenesis in African and South American Weakly Electric Fishes".PLOS ONE.7(5): e36287.Bibcode:2012PLoSO...736287L.doi:10.1371/journal.pone.0036287.PMC3351409.PMID22606250.
- ^Roberts, M.B.V. (1986).Biology: A Functional Approach.Nelson Thornes. p. 274.ISBN978-0-17-448019-8.Archivedfrom the original on 12 September 2016.
- ^Kozmik, Z; Ruzickova, J; Jonasova, K; Matsumoto, Y.; Vopalensky, P.; Kozmikova, I.; Strnad, H.; Kawamura, S.; Piatigorsky, J.; Paces, V.; Vlcek, C. (1 July 2008)."From the Cover: Assembly of the cnidarian camera-type eye from vertebrate-like components".Proceedings of the National Academy of Sciences.105(26): 8989–8993.Bibcode:2008PNAS..105.8989K.doi:10.1073/pnas.0800388105.PMC2449352.PMID18577593.
- ^"Plant and Animal Evolution".University of Waikato.Archivedfrom the original on 18 March 2017.Retrieved10 January2017.
- ^Ben-Hamo, Miriam; Muñoz-Garcia, Agustí; Larrain, Paloma; Pinshow, Berry; Korine, Carmi; Williams, Joseph B. (June 2016)."The cutaneous lipid composition of bat wing and tail membranes: a case of convergent evolution with birds".Proc. R. Soc. B.283(1833): 20160636.doi:10.1098/rspb.2016.0636.PMC4936036.PMID27335420.
- ^Alexander, David E. (2015).On the Wing: Insects, Pterosaurs, Birds, Bats and the Evolution of Animal Flight.Oxford University Press. p. 28.ISBN978-0-19-999679-7.Archivedfrom the original on 14 February 2017.Retrieved21 January2017.
- ^"Analogy: Squirrels and Sugar Gliders".University of California Berkeley.Archivedfrom the original on 27 January 2017.Retrieved10 January2017.
- ^ Herrera, Carlos M. (1992). "Activity pattern and thermal biology of a day-flying hawkmoth (Macroglossum stellatarum) under Mediterranean summer conditions ".Ecological Entomology.17(1): 52–56.Bibcode:1992EcoEn..17...52H.doi:10.1111/j.1365-2311.1992.tb01038.x.hdl:10261/44693.S2CID85320151.
- ^Krenn, Harald W.; Plant, John D.; Szucsich, Nikolaus U. (2005). "Mouthparts of flower-visiting insects".Arthropod Structure & Development.34(1): 1–40.doi:10.1016/j.asd.2004.10.002.
- ^Bauder, Julia A.S.; Lieskonig, Nora R.; Krenn, Harald W. (2011)."The extremely long-tongued Neotropical butterfly Eurybia lycisca (Riodinidae): Proboscis morphology and flower handling".Arthropod Structure & Development.40(2): 122–7.Bibcode:2011ArtSD..40..122B.doi:10.1016/j.asd.2010.11.002.PMC3062012.PMID21115131.
- ^Wilhelmi, Andreas P.; Krenn, Harald W. (2012). "Elongated mouthparts of nectar-feeding Meloidae (Coleoptera)".Zoomorphology.131(4): 325–37.doi:10.1007/s00435-012-0162-3.S2CID9194699.
- ^"When is a thumb a thumb?".Understanding Evolution.Archivedfrom the original on 16 October 2015.Retrieved14 August2015.
- ^abcdeEdwards, M.; et al. (2010)."Association of the OCA2 Polymorphism His615Arg with Melanin Content in East Asian Populations: Further Evidence of Convergent Evolution of Skin Pigmentation".PLOS Genetics.6(3): e1000867.doi:10.1371/journal.pgen.1000867.PMC2832666.PMID20221248.
- ^Meyer, W. K.; et al. (2013)."The convergent evolution of blue iris pigmentation in primates took distinct molecular paths".American Journal of Physical Anthropology.151(3): 398–407.doi:10.1002/ajpa.22280.PMC3746105.PMID23640739.
- ^abPoppenwimer, Tyler; Mayrose, Itay; DeMalach, Niv (December 2023)."Revising the global biogeography of annual and perennial plants".Nature.624(7990): 109–114.arXiv:2304.13101.Bibcode:2023Natur.624..109P.doi:10.1038/s41586-023-06644-x.ISSN1476-4687.PMC10830411.PMID37938778.S2CID260332117.
- ^Friedman, Jannice (2 November 2020)."The Evolution of Annual and Perennial Plant Life Histories: Ecological Correlates and Genetic Mechanisms".Annual Review of Ecology, Evolution, and Systematics.51(1): 461–481.doi:10.1146/annurev-ecolsys-110218-024638.ISSN1543-592X.S2CID225237602.
- ^Hjertaas, Ane C.; Preston, Jill C.; Kainulainen, Kent; Humphreys, Aelys M.; Fjellheim, Siri (2023)."Convergent evolution of the annual life history syndrome from perennial ancestors".Frontiers in Plant Science.13.doi:10.3389/fpls.2022.1048656.ISSN1664-462X.PMC9846227.PMID36684797.
- ^Boyko, James D.; Hagen, Eric R.; Beaulieu, Jeremy M.; Vasconcelos, Thais (November 2023)."The evolutionary responses of life-history strategies to climatic variability in flowering plants".New Phytologist.240(4): 1587–1600.doi:10.1111/nph.18971.ISSN0028-646X.PMID37194450.
- ^Williams, B. P.; Johnston, I. G.; Covshoff, S.; Hibberd, J. M. (September 2013)."Phenotypic landscape inference reveals multiple evolutionary paths to C4 photosynthesis".eLife.2:e00961.doi:10.7554/eLife.00961.PMC3786385.PMID24082995.
- ^Osborne, C. P.;Beerling, D. J.(2006)."Nature's green revolution: the remarkable evolutionary rise of C4plants ".Philosophical Transactions of the Royal Society B: Biological Sciences.361(1465): 173–194.doi:10.1098/rstb.2005.1737.PMC1626541.PMID16553316.
- ^Sage, Rowan; Russell Monson (1999)."16".C4 Plant Biology.Elsevier. pp. 551–580.ISBN978-0-12-614440-6.
- ^Zhu, X. G.; Long, S. P.; Ort, D. R. (2008)."What is the maximum efficiency with which photosynthesis can convert solar energy into biomass?".Current Opinion in Biotechnology.19(2): 153–159.doi:10.1016/j.copbio.2008.02.004.PMID18374559.Archivedfrom the original on 1 April 2019.Retrieved29 December2018.
- ^Sage, Rowan; Russell Monson (1999)."7".C4 Plant Biology.Elsevier. pp. 228–229.ISBN978-0-12-614440-6.
- ^Kadereit, G.; Borsch, T.; Weising, K.; Freitag, H (2003). "Phylogeny of Amaranthaceae and Chenopodiaceae and the Evolution of C4Photosynthesis ".International Journal of Plant Sciences.164(6): 959–86.doi:10.1086/378649.S2CID83564261.
- ^Ireland, Hilary, S.; et al. (2013)."Apple SEPALLATA1/2 -like genes control fruit flesh development and ripening".The Plant Journal.73(6): 1044–1056.doi:10.1111/tpj.12094.PMID23236986.
{{cite journal}}:CS1 maint: multiple names: authors list (link) - ^Heuvelink, Ep (2005).Tomatoes.CABI. p. 72.ISBN978-1-84593-149-0.Archivedfrom the original on 1 April 2019.Retrieved17 December2016.
- ^Lorts, C.; Briggeman, T.; Sang, T. (2008)."Evolution of fruit types and seed dispersal: A phylogenetic and ecological snapshot"(PDF).Journal of Systematics and Evolution.46(3): 396–404.doi:10.3724/SP.J.1002.2008.08039(inactive 1 November 2024). Archived fromthe original(PDF)on 18 July 2013.
{{cite journal}}:CS1 maint: DOI inactive as of November 2024 (link) - ^Lengyel, S.; Gove, A. D.; Latimer, A. M.; Majer, J. D.; Dunn, R. R. (2010). "Convergent evolution of seed dispersal by ants, and phylogeny and biogeography in flowering plants: a global survey".Perspectives in Plant Ecology, Evolution and Systematics.12(1): 43–55.Bibcode:2010PPEES..12...43L.doi:10.1016/j.ppees.2009.08.001.
- ^Fukushima, K; Fang, X; et al. (2017)."Genome of the pitcher plant Cephalotus reveals genetic changes associated with carnivory".Nature Ecology & Evolution.1(3): 0059.Bibcode:2017NatEE...1...59F.doi:10.1038/s41559-016-0059.PMID28812732.
- ^abcStayton, C. Tristan (2015). "The definition, recognition, and interpretation of convergent evolution, and two new measures for quantifying and assessing the significance of convergence".Evolution.69(8): 2140–2153.doi:10.1111/evo.12729.PMID26177938.S2CID3161530.
- ^Stayton, C. Tristan (2008). "Is convergence surprising? An examination of the frequency of convergence in simulated datasets".Journal of Theoretical Biology.252(1): 1–14.Bibcode:2008JThBi.252....1S.doi:10.1016/j.jtbi.2008.01.008.PMID18321532.
- ^Muschick, Moritz; Indermaur, Adrian; Salzburger, Walter (2012)."Convergent Evolution within an Adaptive Radiation of Cichlid Fishes".Current Biology.22(24): 2362–2368.Bibcode:2012CBio...22.2362M.doi:10.1016/j.cub.2012.10.048.PMID23159601.
- ^Arbuckle, Kevin; Bennett, Cheryl M.; Speed, Michael P. (July 2014)."A simple measure of the strength of convergent evolution".Methods in Ecology and Evolution.5(7): 685–693.Bibcode:2014MEcEv...5..685A.doi:10.1111/2041-210X.12195.
- ^Ingram, Travis; Mahler, D. Luke (1 May 2013)."SURFACE: detecting convergent evolution from comparative data by fitting Ornstein-Uhlenbeck models with stepwise Akaike Information Criterion".Methods in Ecology and Evolution.4(5): 416–425.Bibcode:2013MEcEv...4..416I.doi:10.1111/2041-210X.12034.S2CID86382470.
Further reading
edit- Losos, Jonathan B. (2017).Improbable Destinies: Fate, Chance, and the Future of Evolution.Riverhead Books.ISBN978-0399184925.
External links
edit Media related toConvergent evolutionat Wikimedia Commons
Media related toConvergent evolutionat Wikimedia Commons