
Bennettitales
| Bennettitales | |
|---|---|

| |
| Restoration of a member ofWilliamsoniaceaebyThérèse Ekblom | |

| |
| Life restoration of"Williamsonia" sewardianafrom the Early Cretaceous of India, which may represent an early member ofCycadeoidaceae | |
| Scientific classification | |
| Kingdom: | Plantae |
| Clade: | Tracheophytes |
| Clade: | Spermatophytes |
| Order: | †Bennettitales Engler, 1892 |
| Families | |
Bennettitales(also known ascycadeoids) is an extinctorderofseed plantsthat first appeared in thePermianperiod and became extinct in most areas toward the end of theCretaceous.Bennettitales were amongst the most common seed plants of theMesozoic,and had morphologies includingshrubandcycad-like forms. The foliage of bennettitaleans is superficially nearly indistinguishable from that of cycads, but they are distinguished from cycads by their more complex flower-like reproductive organs, at least some of which were likelypollinated by insects.[1]
Although certainlygymnospermssensu lato(cone-bearing seed plants), the relationships of bennettitaleans to other seed plants is debated. Their general resemblance to cycads is contradicted by numerous more subtle features of their reproductive systems and leaf structure. Some authors have linked bennettitaleans toangiosperms(flowering plants) andgnetophytes(a rare and unusual group of modern gymnosperms), forming a broader group known asAnthophyta.Molecular data contradicts this, with gnetophytes found to be much more genetically similar toconifers.The exact position of Bennettitales remains uncertain.
Description[edit]


Bennettitales are divided into two families,CycadeoidaceaeandWilliamsoniaceae,which have distinctgrowth habits.Cycadeoidaceae had stout,cycad-like trunks with bisporangiate (containing bothmegasporesandmicrospores)strobili(cones) serving as theirreproductivestructures. Williamsoniaceae either had bisporangiate or monosporangiate cones, and distinctly slender and branching woody trunks.[1]The Williamsoniaceae grew as woody shrubs with adivaricatebranching habit, similar to that ofBanksia.[2]It has been suggested that Williamsoniaceae are aparaphyletic(not containing all descendants of a common ancestor) assemblage of all Bennettitales that do not belong to the Cycadeoidaceae.[2]
Foliage[edit]
In general, bennettitalean leaves are attached to the stem with a helical (corkscrew) arrangement. Some leaves (most species ofNilssoniopteris,etc.) are narrow, solitary blades with a smooth-edged ( "entire" ) margin.[3]Most leaf morphotypes (Pterophyllum,Ptilophyllum,Zamites,Otozamites,etc.) arepinnate(feather-shaped), with many small leaf segments attached to a central shaft. Others (Anomozamites,a few species ofNilssoniopteris) are incompletely pinnate (sawtooth-shaped) and transitional between these two end members. One unusual leaf form,Eoginkgoites,even approaches apalmateappearance similar to early species ofGinkgo.[4]
The foliage of bennettitaleans resembles that of cycads to such an extent that the foliage of the two groups cannot be reliably distinguished based on gross morphology alone. However, fossil foliage which preserves thecuticlecan be assigned to either group with confidence. Thestomataof bennettitaleans are described assyndetocheilic.This means that the main pairedguard cellsdevelop from the same mother cells as the subsidiary cells which surround them. This contrasts with thehaplocheilicstomata of cycads and conifers. In haplocheilic stomata, the ring of subsidiary cells are not derived from the same original structures as the guard cells. This fundamental difference is the main way to differentiate bennettitalean and cycad foliage.[5]
Cones and seeds[edit]


Like other gymnosperms, bennettitalean reproductive inflorescences come in the form ofcones,which producepollenandovules(unfertilized seeds). The cones have a thick centralreceptaclesurrounded by simple, helically-arranged fertile and infertile structures. Tissue at the base of the cone forms layers of scale-like or petal-likebractsto protect the radiating inner structures. Some authors refer to bennettitalean cones as "flowers", though they are not equivalent to trueangiospermflowers. Pollen is often enclosed in pairedsynangia(pollen sacs). The synangia lie on the adaxial (inner) edge of pollen-bearing leaf-like structures known asmicrosporophylls.This contrasts with cycads, all of which lack discrete synangia and bear pollen on the abaxial (outer) surface of their microsporophylls.[6]
Many bennettitaleans arebisporangiate,where the pollen and ovules are hosted on the same (bisexual or hermaphrodite) cone. Cavities filled with curved synangia-bearing microsporophylls are encased by thin radiating structures, including thick, infertileinterseminal scalesand fertile sporophylls with ovules at their tips. The presence of ovules at the tips of sporophylls, rather than the tips of stems, is a major difference between the cones of bennettitaleans and gnetophytes. As the cone is fertilized and matures, the microsporophylls wither away and the ovules transform into seeds.[6]
Most bennettitaleans in the familyWilliamsoniaceaeare insteadmonosporangiate,with separate pollen and ovule-producing (unisexual) cones on the same plant. The ovule-producing (female) cones (Williamsonia,etc.) are similar to mature bisporangiate cones, with interseminal scales and ovule-tipped sporophylls enclosed by bracts. Pollen-producing (male) cones (Weltrichia,etc.), on the other hand, feature an exposed crown of tapering microsporophylls with adaxial rows of synangia. The microsporophylls may host a single linear row of paired synangia, or instead synangia arranged in a pinnate (feather-shaped) pattern.[6]
Seedsare dicotyledonous (possess twoembryonic leaves), with a central embryo surrounded by three layers: the thin megagametophyte, the slightly thickernucellus,and the protectiveintegument.The upper tip of the seed is tapered and opens through a thin and often extendedmicropyle.A long, narrow micropyle extending out of the seed is superficially similar to the condition in livinggnetophytes.Once the seed is fertilized, the micropyle is sealed by a plug-shaped extension of the nucellus. Unlike living gymnosperms, the tip of the nucellus lacks a pollen chamber (receptacle for stored pollen). The integument is dense and thick, with many layers of differentiated cells. This contrasts with the thin, biseriate (two cell-layer) integument of gnetophytes. Bennettitaleans also lack another gnetophyte-like trait: a sheath of fused bracteoles enveloping the seed. Most integument cells are not unusual in size or shape. However, near the micropyle the innermost layer of integument cells become radially-oriented and elongated, partially closing in on the micropyle. The nucellus and integument are unfused above thechalaza(base of the seed), unlike cycads or gnetophytes, where the layers are fused for much of their height.[6]
Cycadeoidaceans have been suggested to have been self-pollinating, with their stems and cones buried underground,[1][7]although it has alternatively been proposed that they were pollinated by beetles.[8]The flower-like williamsoniacean male reproductive structureWeltrichiais associated with the female reproductive structureWilliamsonia,though it is uncertain whether the parent plants weremonoecious(male and female reproductive structures being present on the same plant) ordioecious(where each plant has only one gender of reproductive organ).Weltrichiawas likely primarilywind-pollinated,with some species possibly pollinated by beetles.[9]
Several groups of Jurassic and Early Cretaceous insects possessed a longproboscis,and it has been suggested that they fed onnectarproduced by bennettitalean reproductive structures, such as the bisexual williamsoniacean reproductive structureWilliamsoniella,which had a long, narrow central receptacle which was likely otherwise inaccessible.[10]Early Cretaceous bennettitalean pollen has been found directly associated with a proboscis bearing fly belonging to the extinct familyZhangsolvidae,providing evidence that this family acted as pollinators for the group.[11]The interseminal scales of Bennettitales ovulate cones may have become fleshy at maturity, which could have potentially made then attractive to wild animals that served as seed dispersers.[12]
Taxonomy[edit]
History of discovery[edit]
The Cycadeoideaceae (originally "Cycadeoideae" ) were named by English geologistWilliam Bucklandin 1828, from fossil trunks found in Jurassic strata on theIsle of Portland,England, which Buckland gave thegenusnameCycadeoidea.Buckland provided a description of the family and two species, but failed to give a description of the genus, which has led to Buckland's description of the family being considered invalid by modern taxonomic standards.[13]In publications in 1870, Scottish botanistWilliam Carruthersand English paleobotanistWilliam Crawford Williamsondescribed the first known reproductive organs of the Bennettitales from Jurassic strata ofYorkshireand Jurassic-Cretaceous strata of theIsle of Wightand the Isle of Portland.[14][15][16]Caruthers was the first to recognise that Bennettitales had distinct differences from cycads, and established the tribes "Williamsonieae" and "Bennettiteae",[16]with the latter being named after the genusBennettitesnamed by Caruthers in the same publication,the name being in honour of British botanistJohn Joseph Bennett.[14][17]The order Bennettitales was erected by German botanistAdolf Englerin 1892, who recognised the group as separate from the Cycadales.[18]
Relationships to other seed plants[edit]
TheAnthophytehypothesis erected by Arber and Parking in 1907[19]posited thatangiospermsarose from Bennettitales, as suggested by the wood-like structures and rudimentaryflowers.[1]Based on morphological data, however, Bennettitales were classified as amonophyletic groupwhen paired withGnetales.[20]a study in 2006 suggested that Bennettitales, Angiosperms, andGigantopteridalesform acladebased on the presence ofoleanane.[21]Molecular evidence has consistently contradicted the Anthophyte hypothesis, finding that Angiosperms are the sister group to all living gymnosperms, including Gnetales.[22]Some authors have suggested due to similarities between their seed coats, Bennettitales form a clade with the gymnosperm orders ofGnetalesandErdtmanithecales,dubbed the "BEG group".[23]However, this proposal has been contested by other authors, who contend that these similarities are only superficial and do not indicate a close relationship.[24]A 2017 phylogeny based on molecular signatures of fossilised cuticles found that Bennettitales were more closely related to theGinkgo+Cycads clade than conifers, and were closely related toNilssoniaandPtilozamites.[25]
Evolutionary history[edit]
The oldest confirmed fossils of bennettitaleans are leaves ofNilssoniopteris shanxiensis,a species from the upper part of the UpperShihhotse FormationinShanxi Province,China.[5]This strata is dated to the earlyKungurianstage of the earlyPermian(Cisuralian), around 281 million years ago.[26]Supposed Carboniferous-Permian records ofPterophyllumdo not have conclusive bennettitalean affinities or have been reinterpreted as cycad foliage in the form genusPseudoctenis.[27]True Permian records of benettitalean leaves are rare; outside of the Shihhotse Formation they are only found in theLate Permian(likelyChanghsingian)-ageUmm Irna FormationinJordan.[5]This formation is notable for the early occurrence of other Mesozoic-style flora, including the earliest records ofcorystospermaleanfoliage (Dicroidium).[28]The order Fredlindiales (containing the genusFredlindia) from the Late Triassic of Gondwana appears to be closely related to Bennettitales, but differs from it in some aspects of its reproductive organs.[16]
The bennettitalean fossil record reappeared in theMiddle Triassic,and williamsoniaceans became globally distributed by the end of the period.[29][5]The oldest bennettitalean reproductive structures are smallWilliamsonia"flowers" from the Middle TriassicEsk FormationofAustralia.[16]While Williamsoniaceae had a global distribution, Cycadeoidaceae appear to have been primarily confined to the western parts ofLaurasia,and are primarily known from the Cretaceous.[16]Bennettitales were widespread and abundant during the Jurassic and Early Cretaceous, however Bennettitales severely declined during the Late Cretaceous, coincident with the rise offlowering plants,being mostly extinct by the end of the period, with the final known remains from the Northern Hemisphere being found in the polar latitudeKakanaut FormationinChukotka,Russia, dating to theMaastrichtian,assignable toPterophyllum.[30]A possible late record has been reported from the earlyOligoceneof eastern Australia andTasmania,assignable to the genusPtilophyllum,but no cuticle was preserved, making the referral inconclusive.[31]
Subgroups[edit]
- Cycadolepis(scales or bracts, unplaced in family)
- Haitingeria?(pollen organ?)
- Lunzia(pollen organ, unplaced in family)
- Leguminanthus?(pollen organ?)
- Leuthardtia?(pollen organ?)
- Westersheimia(ovulate organ, unplaced in family)
- Anthrophyopsis?[32](leaf)
- FamilyCycadeoidaceae
- FamilyWilliamsoniaceae
- Anomozamites(leaf)
- Bennetticarpus(female seed cone)
- Bennettistemon(male pollen organ)
- Bucklandia(axes)
- Eoginkgoites(leaf)
- Ischnophyton
- Kimuriella(whole plant)
- Nilssoniopteris(leaf)
- Otozamites(leaf)
- Pterophyllum(leaf)
- Ptilophyllum(leaf)
- Vardekloeftia(female seed cone)
- Weltrichia(male pollen organ)
- Wielandiella(whole plant)
- Williamsonia(female seed cones)
- Williamsoniella(bisexual reproductive structure)
- Zamites(leaf, in partim)
- Haitingeria(pollen organ)
Bennettitales is typically considered the sole order in theclassBennettitopsidaEngler (1897)orCycadeoideopsidaScott (1923).Most paleobotanists prefer the two families as used here, though some authors, such as Anderson & Anderson (2007), classify the order via a larger number of families.[33]Anderson & Anderson also classified the ordersFredlindialesAnderson & Anderson (2003)[34]andPentoxylalesPilger & Melchior (1954)within Bennettitopsida.[33]
Gallery[edit]
-
 Fossil leaf ofZamites mariposanafrom theJurassic.
Fossil leaf ofZamites mariposanafrom theJurassic. -
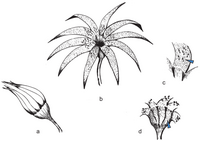 Restoration ofWeltrichia magnafrom the Jurassic of Mexico
Restoration ofWeltrichia magnafrom the Jurassic of Mexico -
 Otozamitesleaf
Otozamitesleaf -
 Leaf ofAnomozamites
Leaf ofAnomozamites -
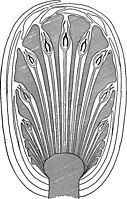 Seed bearingstrobilusofBennettites
Seed bearingstrobilusofBennettites -
 Trunk ofCycadeoidea
Trunk ofCycadeoidea -
 Fossil ofPtilophyllum grandifolium
Fossil ofPtilophyllum grandifolium







References[edit]
- ^abcdPeñalver, Enrique; Arillo, Antonio; Pérez-de la Fuente, Ricardo; Riccio, Mark L.; Delclòs, Xavier; Barrón, Eduardo; Grimaldi, David A. (2015)."Long-Proboscid Flies as Pollinators of Cretaceous Gymnosperms".Current Biology.25(14): 1917–1923.doi:10.1016/j.cub.2015.05.062.PMID26166781.S2CID13022302.
- ^abPott, Christian; McLoughlin, Stephen (2014-06-01)."Divaricate growth habit in Williamsoniaceae (Bennettitales): unravelling the ecology of a key Mesozoic plant group".Palaeobiodiversity and Palaeoenvironments.94(2): 307–325.doi:10.1007/s12549-014-0157-9.S2CID84440045.
- ^Ray, M.M.; Rothwell, G.W.; Stockey, R.A. (September 2014)."Anatomically Preserved Early Cretaceous Bennettitalean Leaves: Nilssoniopteris corrugata n. sp. from Vancouver Island, Canada".Journal of Paleontology.88(5): 1085–93.doi:10.1017/S002233600005767X.S2CID232349931.
- ^Pott, Christian; Axsmith, Brian J. (February 2015)."Williamsonia carolinensis sp. nov. and Associated Eoginkgoites Foliage from the Upper Triassic Pekin Formation, North Carolina: Implications for Early Evolution in the Williamsoniaceae (Bennettitales)".International Journal of Plant Sciences.176(2): 174–185.doi:10.1086/679471.S2CID44559347.
- ^abcdBlomenkemper, Patrick; Bäumer, Robert; Backer, Malte; Abu Hamad, Abdalla; Wang, Jun; Kerp, Hans; Bomfleur, Benjamin (2021)."Bennettitalean Leaves From the Permian of Equatorial Pangea—The Early Radiation of an Iconic Mesozoic Gymnosperm Group".Frontiers in Earth Science.9:162.Bibcode:2021FrEaS...9..162B.doi:10.3389/feart.2021.652699.
- ^abcdRothwell, Gar W.; Crepet, William L.; Stockey, Ruth A. (2009)."Is the anthophyte hypothesis alive and well? New evidence from the reproductive structures of Bennettitales".American Journal of Botany.96(1): 296–322.doi:10.3732/ajb.0800209.PMID21628190.
- ^Osborn, J.M.; Taylor, M.L. (2010). "Pollen and coprolite structure in Cycadeoidea (Bennettitales): implications for understanding pollination and mating systems in Mesozoic cycadeoids".Plants in deep Mesozoic time: morphological innovations, phylogeny, and ecosystems.Bloomington, IN: Indiana University Press. pp. 34–49.ISBN978-0-253-00199-3.
- ^Frame, Dawn; Gottsberger, Gerhard (21 April 2023)."Diverse sexual strategies in fossil gymnosperms: pollination in the Bennettitales revisited".Phyton.62/63.
- ^Popa, Mihai Emilian (2019-02-27)."Review of the bennettitalean genus Weltrichia".Journal of Palaeogeography.8(1): 12.Bibcode:2019JPalg...8...12P.doi:10.1186/s42501-019-0023-9.S2CID85458053.
- ^Khramov, Alexander V.; Lukashevich, Elena D. (July 2019). "A Jurassic dipteran pollinator with an extremely long proboscis".Gondwana Research.71:210–215.Bibcode:2019GondR..71..210K.doi:10.1016/j.gr.2019.02.004.S2CID134847380.
- ^Peñalver E, Arillo A, Pérez-de la Fuente R, Riccio ML, Delclòs X, Barrón E, Grimaldi DA (July 2015)."Long-Proboscid Flies as Pollinators of Cretaceous Gymnosperms".Current Biology.25(14): 1917–23.doi:10.1016/j.cub.2015.05.062.PMID26166781.
- ^Friis, Else Marie; Pedersen, Kaj Raunsgaard; Crane, Peter R., eds. (2011),"Angiosperms in context: extant and fossil seed plants",Early Flowers and Angiosperm Evolution,Cambridge: Cambridge University Press, pp. 101–140,ISBN978-0-521-59283-3,retrieved2023-03-20
- ^Doweld, Alexander B. (2016-05-03)."The nomenclature of Cycadeoidea (fossil Spermatophyta: Cycadeoideopsida)".Taxon.65(2): 372–379.doi:10.12705/652.16.
- ^abCarruthers, William (May 1870)."XVIII. On Fossil Cycadean Stems from the Secondary Rocks of Britain".Transactions of the Linnean Society of London.26(4): 675–708.doi:10.1111/j.1096-3642.1870.tb00201.x.
- ^Williamson, W. C. (May 1870)."XVII. Contributions towards the History of Zamia Gigas, Lindl. & Hutt.*".Transactions of the Linnean Society of London.26(4): 663–674.doi:10.1111/j.1096-3642.1870.tb00200.x.
- ^abcdeMcLoughlin, Stephen; Pott, Christian; Sobbe, Ian H. (2018-03-01)."The diversity of Australian Mesozoic bennettitopsid reproductive organs"(PDF).Palaeobiodiversity and Palaeoenvironments.98(1): 71–95.doi:10.1007/s12549-017-0286-z.S2CID135237376.
- ^Britten, James; Boulger, George Simonds (1893).A biographical index of British and Irish botanists.London: West, Newman & co. p. 14.doi:10.5962/bhl.title.56947.
- ^Engler, H.G.A. 1892.Syllabus der Vorlesungen über specielle und medicinisch-pharmaceutische Botanik.xxiii + 184 pp. Gebr. Borntraeger, Berlin
- ^Arber, E.N.; Parkin, J. (July 1907)."On the Origin of Angiosperms".Botanical Journal of the Linnean Society.38(263): 29–80.doi:10.1111/j.1095-8339.1907.tb01074.x.
- ^Crane, P.R. (September 1985). "Phylogenetic relationships in seed plants".Cladistics.1(4): 329–348.doi:10.1111/j.1096-0031.1985.tb00432.x.PMID34965681.S2CID85709030.
- ^Taylor, David Winship; Li, Hongqi; Dahl, Jeremy; Fago, Fred J.; Zinniker, David; Moldowan, J. Michael (March 2006). "Biogeochemical evidence for the presence of the angiosperm molecular fossil oleanane in Paleozoic and Mesozoic non-angiospermous fossils".Paleobiology.32(2): 179–90.doi:10.1666/0094-8373(2006)32[179:BEFTPO]2.0.CO;2.S2CID83801635.
- ^Ran, Jin-Hua; Shen, Ting-Ting; Wang, Ming-Ming; Wang, Xiao-Quan (June 2018)."Phylogenomics resolves the deep phylogeny of seed plants and indicates partial convergent or homoplastic evolution between Gnetales and angiosperms".Proceedings. Biological Sciences.285(1881): 20181012.doi:10.1098/rspb.2018.1012.PMC6030518.PMID29925623.
- ^Friis, Else Marie; Crane, Peter R.; Pedersen, Kaj Raunsgaard; Bengtson, Stefan; Donoghue, Philip C. J.; Grimm, Guido W.; Stampanoni, Marco (November 2007)."Phase-contrast X-ray microtomography links Cretaceous seeds with Gnetales and Bennettitales".Nature.450(7169): 549–52.Bibcode:2007Natur.450..549F.doi:10.1038/nature06278.PMID18033296.S2CID1198220.
- ^Rothwell, Gar W.; Stockey, Ruth A. (March 2013)."Evolution and Phylogeny of Gnetophytes: Evidence from the Anatomically Preserved Seed Cone Protoephedrites eamesii gen. et sp. nov. and the Seeds of Several Bennettitalean Species".International Journal of Plant Sciences.174(3): 511–529.doi:10.1086/668688.ISSN1058-5893.S2CID84063572.
- ^Vajda, Vivi; Pucetaite, Milda; McLoughlin, Stephen; Engdahl, Anders; Heimdal, Jimmy; Uvdal, Per (August 2017)."Molecular signatures of fossil leaves provide unexpected new evidence for extinct plant relationships".Nature Ecology & Evolution.1(8): 1093–1099.doi:10.1038/s41559-017-0224-5.PMID29046567.S2CID3604369.
- ^Wu, Qiong; Ramezani, Jahandar; Zhang, Hua; Wang, Jun; Zeng, Fangui; Zhang, Yichun; Liu, Feng; Chen, Jun; Cai, Yaofeng; Hou, Zhangshuai; Liu, Chao (2021-02-05)."High-precision U-Pb age constraints on the Permian floral turnovers, paleoclimate change, and tectonics of the North China block".Geology.49(6): 677–681.Bibcode:2021Geo....49..677W.doi:10.1130/G48051.1.S2CID234064909.
- ^Pott, Christian; McLoughlin, Stephen; Lindström, Anna (2009)."Late Palaeozoic Foliage from China Displays Affinities to Cycadales Rather than to Bennettitales Necessitating a Re-Evaluation of the Palaeozoic Pterophyllum Species".Acta Palaeontologica Polonica.55(1): 157–168.doi:10.4202/app.2009.0070.S2CID52253114.
- ^Blomenkemper, Patrick; Kerp, Hans; Abu Hamad, Abdalla; DiMichele, William A.; Bomfleur, Benjamin (December 2018)."A hidden cradle of plant evolution in Permian tropical lowlands".Science.362(6421): 1414–1416.Bibcode:2018Sci...362.1414B.doi:10.1126/science.aau4061.PMID30573628.
- ^Kustatscher, Evelyn; van Konijnenburg-van Cittert, Johanna H.A. (2005)."The Ladinian Flora (Middle Triassic) of the Dolomites: palaeoenvironmental reconstructions and palaeoclimatic considerations"(PDF).Geo.Alp.2:31–51.
- ^Gnilovskaya, Anastasia A.; Golovneva, Lina B. (February 2018)."The Late Cretaceous Pterophyllum (Bennettitales) in the North-East of Russia".Cretaceous Research.82:56–63.doi:10.1016/j.cretres.2017.09.013.
- ^McLoughlin, Stephen; Carpenter, Raymond J.; Pott, Christian (April 2011)."Ptilophyllum muelleri (Ettingsh.) comb. nov. from the Oligocene of Australia: last of the Bennettitales?".International Journal of Plant Sciences.172(4): 574–85.doi:10.1086/658920.S2CID52885618.
- ^Xu, Yuanyuan; Wang, Yongdong; McLoughlin, Stephen (September 2023)."How similar are the venation and cuticular characters of Glossopteris, Sagenopteris and Anthrophyopsis?".Review of Palaeobotany and Palynology.316:104934.doi:10.1016/j.revpalbo.2023.104934.
- ^abAnderson, John M.; Anderson, Heidi M.; Cleal, Chris J. (2007)."Brief history of the gymnosperms: classification, biodiversity, phytogeography and ecology"(PDF).Strelitzia.20:1–280.
- ^Anderson, John M.; Anderson, Heidi M. (2003)."Heyday of the gymnosperms: systematics and biodiversity of the Late Triassic Molteno fructifications".Strelitzia.15:1–308.