











Articles
https://doi.org/10.1038/s41561-018-0133-5
Tall Amazonian forests are less sensitive to
precipitation variability
Francesco Giardina
1,2
, Alexandra G. Konings
3
, Daniel Kennedy
4
, Seyed Hamed Alemohammad
4
,
Rafael S. Oliveira
5,6
, Maria Uriarte
7
and Pierre Gentine
4,8
*
1
Department of Earth and Environmental Engineering, Columbia University, New York, NY, USA.
2
École Polytechnique Fédérale de Lausanne, Lausanne,
Switzerland.
3
Department of Earth System Science, Stanford University, Stanford, CA, USA.
4
Department of Earth and Environmental Engineering,
Columbia University, New York, NY, USA.
5
Departamento de Biologia Vegetal, Universidade Estadual de Campinas, Campinas, Brazil.
6
School of Plant
Biology, University of Western Australia, Perth, Western Australia, Australia.
7
Department of Ecology, Evolution and Environmental Biology, Columbia
University, New York, NY, USA.
8
The Earth Institute, Columbia University, New York, NY, USA. *e-mail: [email protected]
© 2018 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
SUPPLEMENTARY INFORMATION
In the format provided by the authors and unedited.
NATURE GEOSCIENCE | www.nature.com/naturegeoscience
1
Supplementary Material
Modelling study experiment description
Our study includes two simulation experiments with a plant hydraulic model to support the
observational evidence that taller trees are less sensitivity to variability in precipitation. Since it is
difficult to estimate variations in plant hydraulic traits across the Amazon, we do not try to
explicitly model any particular scenario or location. Instead, we employ two simple experiments
designed to isolate – and illustrate – the role of tree height on the sensitivity of photosynthesis to
water availability. Experiment 1 is an extended drydown, repeating the same single day forcing,
sixty times. Forcing data was based on the average diurnal cycle of September, 2001 at the
Caxiuanã flux tower site (Fluxnet BR-Cax site). An abbreviated model description is provided in
the main text (Methods) and the full model code and forcing data are available online at
github.com/djk2120/talltrees. We run the soil-plant-atmosphere continuum model for two cases,
representing a taller and shorter tree. The only parameters that differ between the cases are: tree
height (z), rooting depth (Z
r
), and the water potential at 50% loss of stomatal conductance (p50).
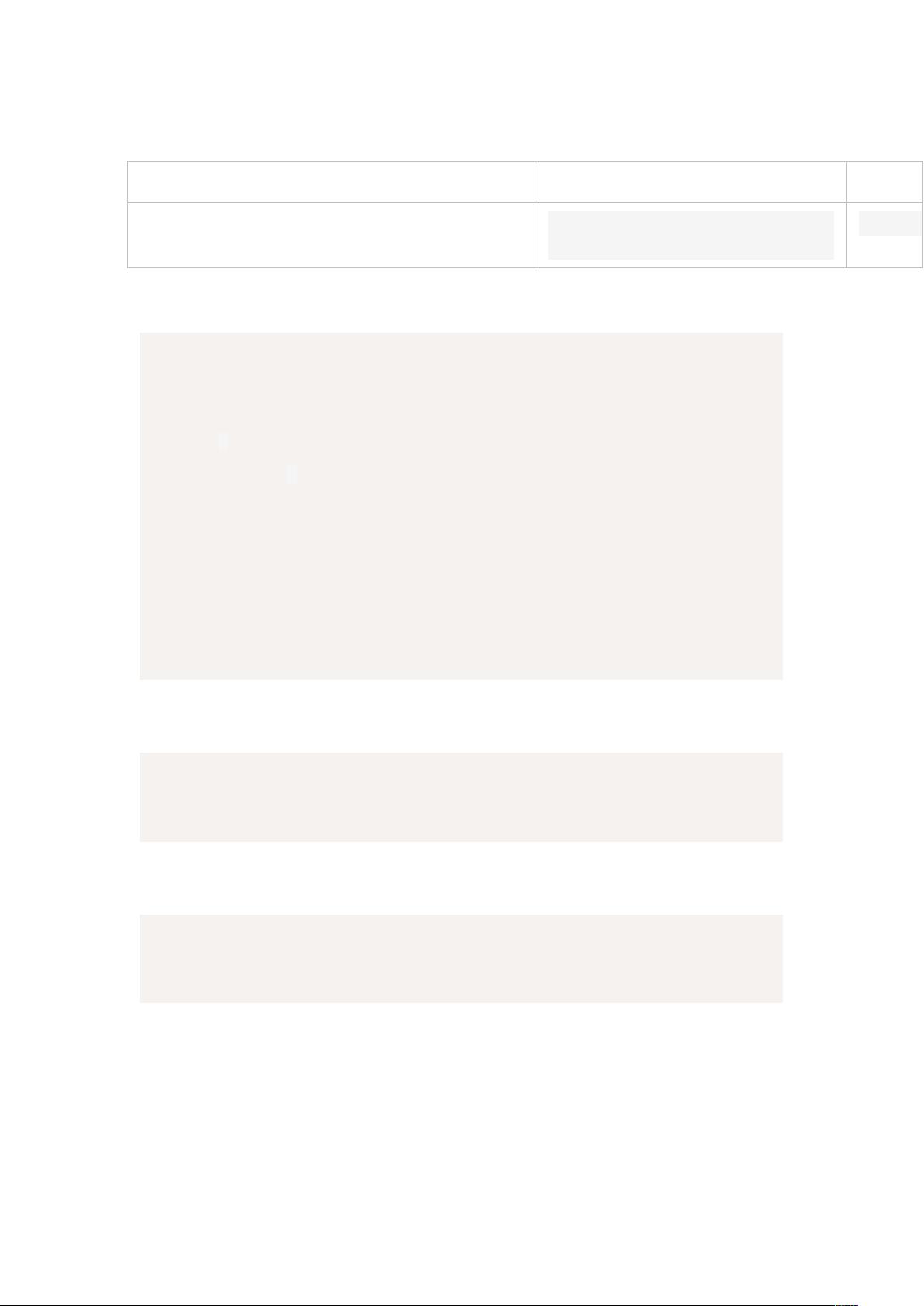
Table S1: Modeling experiment hydraulics
parameters
Units
Short
Tall
z
Tree height
meters
15
40
Kmax
Maximum hydraulic conductivity.
Leaf area basis. Unit tree height (m).
mmol⋅MPa
-1
⋅s
-1
⋅m
-1
220
220
Z
r
Rooting depth
meters
2
3
p1
Xylem vulnerability onset
MPa
-1
-1
p2
Point of 100% loss of xylem
conductivity
MPa
-4
-4
p50
Point of 50% loss of stomatal
conductance
MPa
-1.5
-2.5
a
Shape fitting parameter for loss of
stomatal conductance
(-)
6
1
6
1
1
Value from Xu et al. 2016 for seasonally dry tropical forest
Experiment 2 is the same as Experiment 1, but we remove the bucket model for the tall tree
case. Instead, we force the simulation with the soil moisture from the shorter case, in Experiment 1.
This allows us to distinguish the effect of rooting depth from physiological controls.
For Experiment 3, we apply an atmospheric drydown to test the sensitivity of short vs. tall trees
to changes in vapour pressure deficit. Over a 60-day drydown, we decrease relative humidity each
day by a fixed value (ΔRH = -0.9% per day, which corresponds to -38 Pa per day vapour pressure
deficit at midday) relative to the daily forcing from Experiment 1. We use the same hydraulics
parameters as Experiment 1, but a constant soil potential forcing of -0.2 MPa, in order to empahisze
the role of vapour pressure deficit.
Modelling study results
The taller tree case is less sensitive to the drydown. This is true in both experiments as seen in
Supplementary Figure 4b,c. Cumulative photosynthesis on day 60 is 76.4% of day 1 photosynthesis
for the shorter tree case (same for both experiments). In Experiment 1, the taller trees maintained
94.1% of the day 1 photosynthesis. This is largely due to the effect of deeper rooting, as the shorter
trees experienced more negative soil potentials, as low as -1.34 MPa versus only -0.79 for the taller
case (Fig S5a).
Experiment 2 eliminates the effect of deep roots, by using the soil potential forcing of the short
tree for the tall case. Likewise, the parameter values chosen lead to comparable anisohydricity
(defined as the slope of midday leaf water potential to soil water potential), with σ=0.74 for the
short trees and σ=0.72 for the tall trees (Supplementary Figure 5b). In Experiment 2, without deeper
rooting, tall trees are still less sensitive to the drydown than the shorter trees, maintaining 83.1% of

2
the day 1 photosynthesis (Supplementary Figure 4b). This experiment is used to highlight a second
mechanism for tall forest resilience to precipitation variability unrelated to rooting depth and the
anisohydricity metric, where taller trees are less sensitive to drops in soil potential, similary to our
observations (see main text).
Indeed, this can be explained by the change in midday minus predawn leaf water potential
(Δ
𝜓
) induced by the drydown. In the shorter tree case, the potential drop Δ
𝜓
on day 1 is 0.63 MPa
(Supplementary Figure 6a). On day 60, the drop is 0.34 MPa. This represents a loss of 45%
(0.28MPa) of the day 1 potential drop. For the taller tree case, using the same soil potential forcing
(Experiment 2), the day 1 potential drop is -1.55 MPa. This derives from the lower conductance for
the taller tree case, due to a longer xylem path and viscous dissipation (Darcy’s law). The tall tree
day 60 potential drop Δ
𝜓
is -1.24 MPa (Supplementary Figure 6b), representing a loss of 19.7%
(0.31 MPa) of the day 1 potential drop. Though the absolute change in potential drop is comparable
for the tall and short trees (due to the nearly equivalent anisohydricity), this relative change is larger
for the shorter tree. Transpiration supply is proportional to the gradient in water potential, whereby
the effect of drydown is larger for the shorter trees, due to the relatively large loss in potential
gradient.
The taller tree case is more sensitive to atmospheric demand, Supplementary Figures 4a and 7.
Indeed, the taller tree experiences more stress on the first day of the drydown around midday, which
is associated with high transpiration demand due to high VPD and photosynthesis, while soil water
potential has not had the time to change. To further explore this, we carried out another experiment
simulating the atmospheric drydown only, increasing VPD over the course of 60 days (Experiment
3). The midday gross photosynthesis was more sensitive to higher VPD in the taller tree case,
decreasing by 2.58 µmol/m
2
/d as compared to a drop by just 1.45 µmol/m
2
/d in the shorter tree case
(Supplementary Fig. 7). The overall sensitivity for both to VPD is relatively small, because we
prescribed well-watered conditions (constant soil potential of -0.2 MPa), to decompose the soil and
atmospheric drying effects.

3
Supplementary Figures
Supplementary Figure 1 | Variations of SIF sensitivity to VPD and precipitation binned by
tree height (without 2015) and anisohydricity. a,b Partial correlation coefficients for the
sensitivity of mean SIF to mean precipitation (blue line) and maximum VPD (red line), binned by
tree height, without the 2015 El-Niño year (a) and binned by anisohydricity (b). Normalized yearly
values were used for every variable. c, Spatial pattern of anisohydricity in the Amazon. Only
tropical rainforests are shown d, Correlation coefficients between precipitation with max VPD
(light blue line) and mean VPD (dark red line), binned by tree height.

4
Supplementary Figure 2 | Correlation of mean interannual precipitation with canopy
characteristics and precipitation. a, Scatter plot between precipitation and tree age. b, Scatter plot
between precipitation and aboveground biomass. c, Scatter plot between precipitation and
anisohydricity. d, Scatter plot between mean interannual precipitation and the interannual
variability of mean precipitation. Coefficients of determinations are shown in every plot. The
significance of correlation was calculated using a t-test with a significance level of 0.05. All
correlations are significant at p < 0.001.















